

Novel endoribonucleases as central players in various pathways of eukaryotic RNA metabolism
- PMID: 20675404
- PMCID: PMC2924532
- DOI: 10.1261/rna.2237610
Novel endoribonucleases as central players in various pathways of eukaryotic RNA metabolism
Abstract
For a long time it has been assumed that the decay of RNA in eukaryotes is mainly carried out by exoribonucleases, which is in contrast to bacteria, where endoribonucleases are well documented to initiate RNA degradation. In recent years, several as yet unknown endonucleases have been described, which has changed our view on eukaryotic RNA metabolism. Most importantly, it was shown that the primary eukaryotic 3' --> 5' exonuclease, the exosome complex has the ability to endonucleolytically cleave its physiological RNA substrates, and novel endonucleases involved in both nuclear and cytoplasmic RNA surveillance pathways were discovered concurrently. In addition, endoribonucleases responsible for long-known processing steps in the maturation pathways of various RNA classes were recently identified. Moreover, one of the most intensely studied RNA decay pathways--RNAi--is controlled and stimulated by the action of different endonucleases. Furthermore, endoribonucleolytic cleavages executed by various enzymes are also the hallmark of RNA degradation and processing in plant chloroplasts. Finally, multiple context-specific endoribonucleases control qualitative and/or quantitative changes of selected transcripts under particular conditions in different eukaryotic organisms. The aim of this review is to discuss the impact of all of these discoveries on our current understanding of eukaryotic RNA metabolism.
Figures


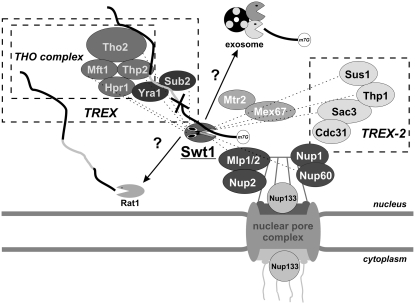

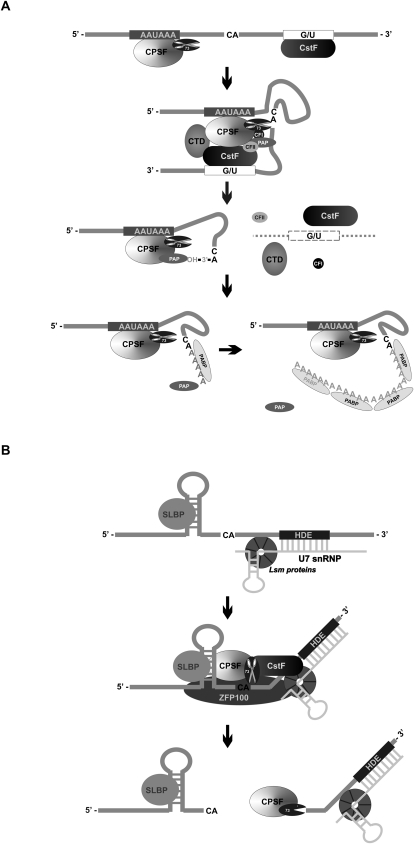

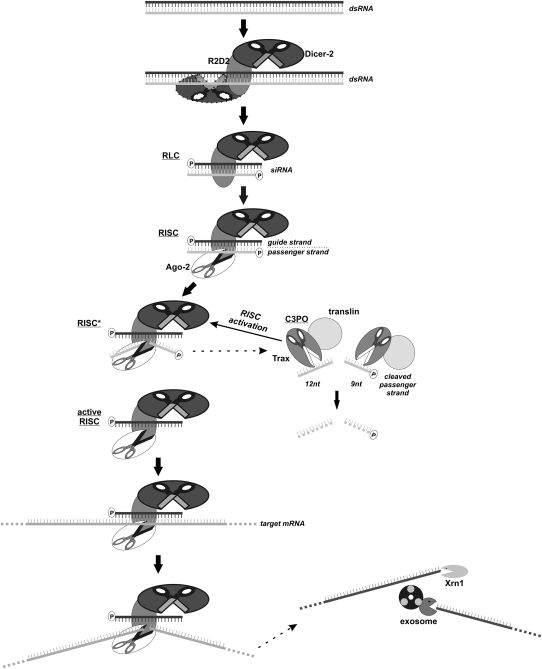
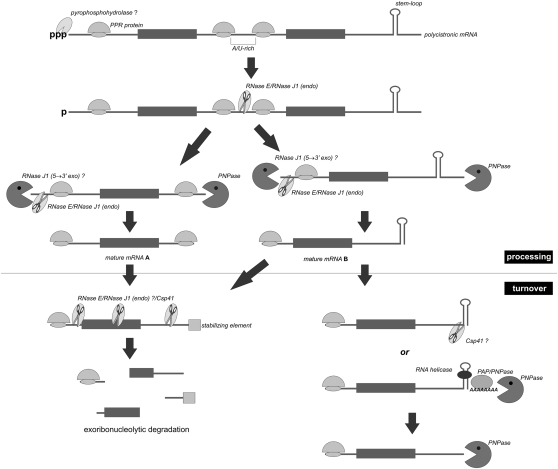


References
-
- Amrani N, Sachs MS, Jacobson A 2006. Early nonsense: mRNA decay solves a translational problem. Nat Rev Mol Cell Biol 7: 415–425 - PubMed
-
- Baillat D, Hakimi MA, Naar AM, Shilatifard A, Cooch N, Shiekhattar R 2005. Integrator, a multiprotein mediator of small nuclear RNA processing, associates with the C-terminal repeat of RNA polymerase II. Cell 123: 265–276 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources