

Nf2/Merlin controls progenitor homeostasis and tumorigenesis in the liver
- PMID: 20675406
- PMCID: PMC2922501
- DOI: 10.1101/gad.1938710
Nf2/Merlin controls progenitor homeostasis and tumorigenesis in the liver
Abstract
The molecular signals that control the maintenance and activation of liver stem/progenitor cells are poorly understood, and the role of liver progenitor cells in hepatic tumorigenesis is unclear. We report here that liver-specific deletion of the neurofibromatosis type 2 (Nf2) tumor suppressor gene in the developing or adult mouse specifically yields a dramatic, progressive expansion of progenitor cells throughout the liver without affecting differentiated hepatocytes. All surviving mice eventually developed both cholangiocellular and hepatocellular carcinoma, suggesting that Nf2(-/-) progenitors can be a cell of origin for these tumors. Despite the suggested link between Nf2 and the Hpo/Wts/Yki signaling pathway in Drosophila, and recent studies linking the corresponding Mst/Lats/Yap pathway to mammalian liver tumorigenesis, our molecular studies suggest that Merlin is not a major regulator of YAP in liver progenitors, and that the overproliferation of Nf2(-/-) liver progenitors is instead driven by aberrant epidermal growth factor receptor (EGFR) activity. Indeed, pharmacologic inhibition of EGFR blocks the proliferation of Nf2(-/-) liver progenitors in vitro and in vivo, consistent with recent studies indicating that the Nf2-encoded protein Merlin can control the abundance and signaling of membrane receptors such as EGFR. Together, our findings uncover a critical role for Nf2/Merlin in controlling homeostasis of the liver stem cell niche.
Figures
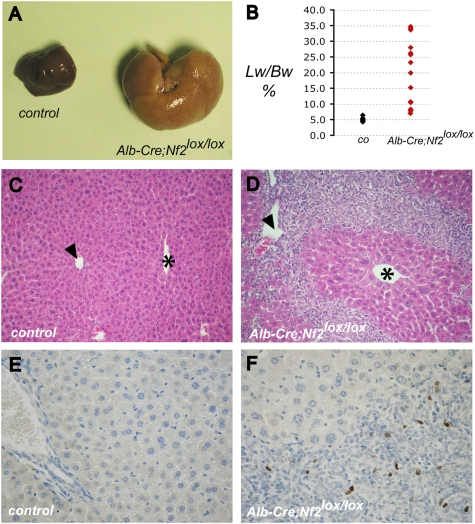
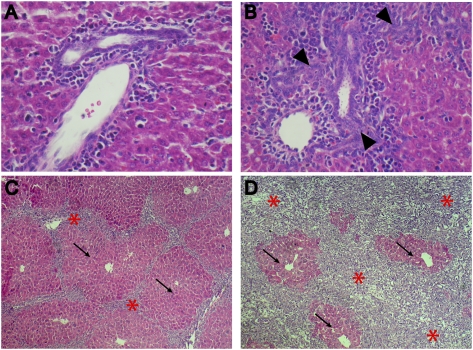
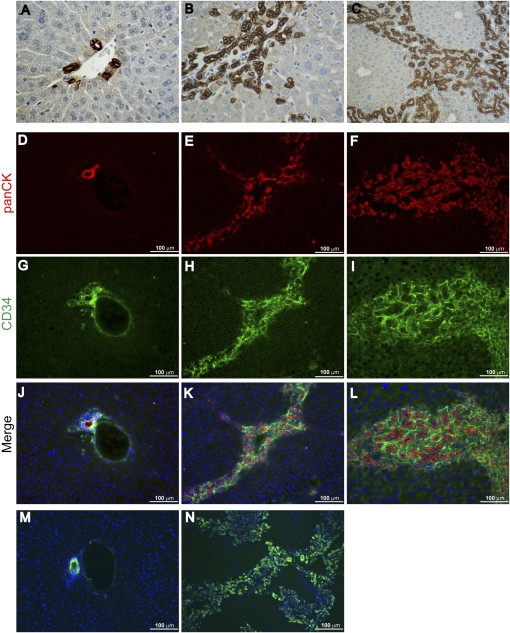
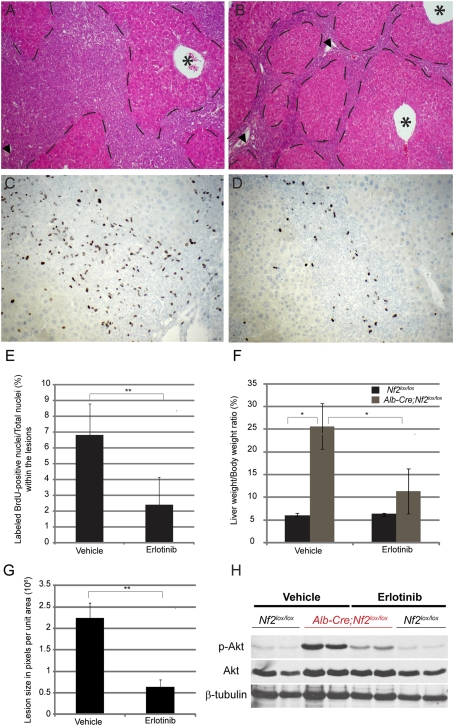
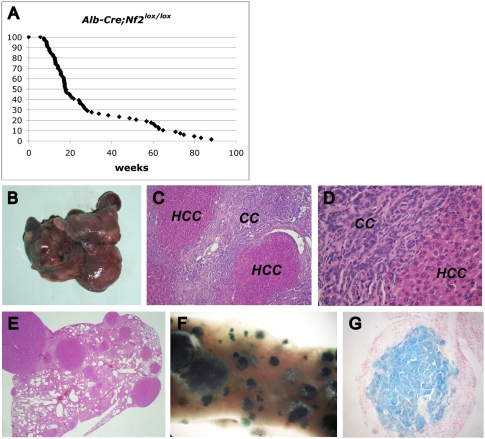
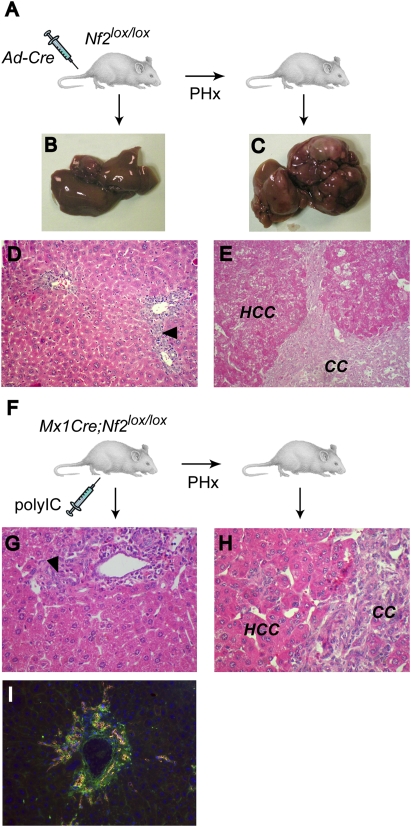
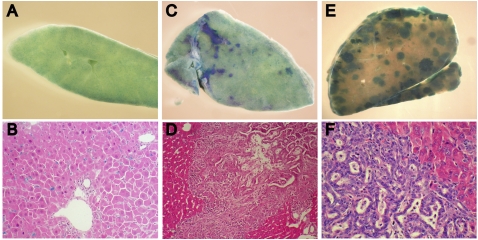
Comment in
-
Merlin in organ size control and tumorigenesis: Hippo versus EGFR?Genes Dev. 2010 Aug 15;24(16):1673-9. doi: 10.1101/gad.1964810. Genes Dev. 2010. PMID: 20713513 Free PMC article.
-
A cellular view of Nf2 in liver homeostasis and tumorigenesis.Dev Cell. 2010 Sep 14;19(3):363-4. doi: 10.1016/j.devcel.2010.08.015. Dev Cell. 2010. PMID: 20833359
-
Tumorigenesis: Merlin, the liver wizard.Nat Rev Cancer. 2010 Oct;10(10):666. doi: 10.1038/nrc2936. Nat Rev Cancer. 2010. PMID: 21080567 No abstract available.
-
Neurofibromatosis type 2/Merlin: sharpening the myth of prometheus.Hepatology. 2011 May;53(5):1767-70. doi: 10.1002/hep.24282. Hepatology. 2011. PMID: 21520182
References
-
- Adams GB, Chabner KT, Alley IR, Olson DP, Szczepiorkowski ZM, Poznansky MC, Kos CH, Pollak MR, Brown EM, Scadden DT 2006. Stem cell engraftment at the endosteal niche is specified by the calcium-sensing receptor. Nature 439: 599–603 - PubMed
-
- Alison MR 2005. Liver stem cells: Implications for hepatocarcinogenesis. Stem Cell Rev 1: 253–260 - PubMed
-
- Baumgartner R, Poernbacher I, Buser N, Hafen E, Stocker H 2010. The WW domain protein Kibra acts upstream of Hippo in Drosophila. Dev Cell 18: 309–316 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous