

The cytoskeleton enhances gene expression in the response to the Harpin elicitor in grapevine
- PMID: 20675535
- PMCID: PMC2935876
- DOI: 10.1093/jxb/erq221
The cytoskeleton enhances gene expression in the response to the Harpin elicitor in grapevine
Abstract
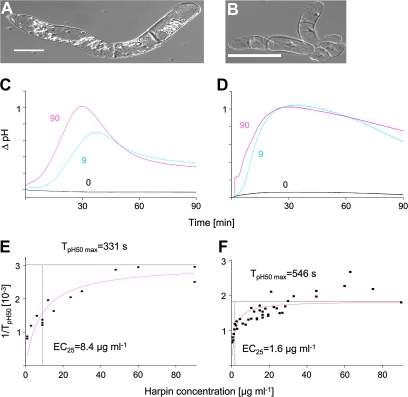
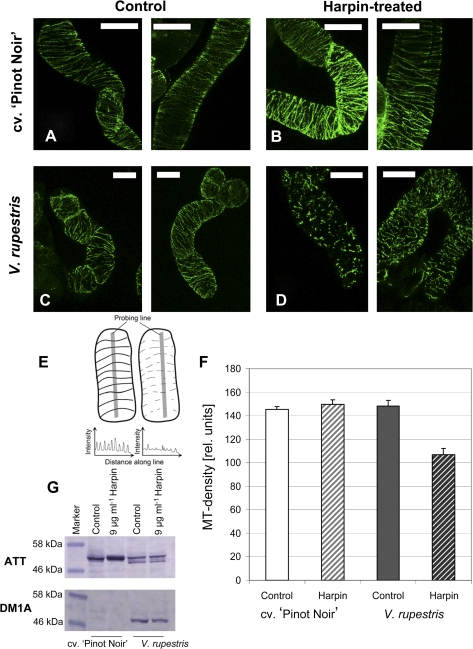
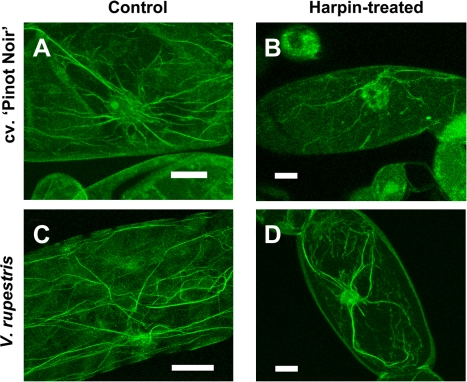
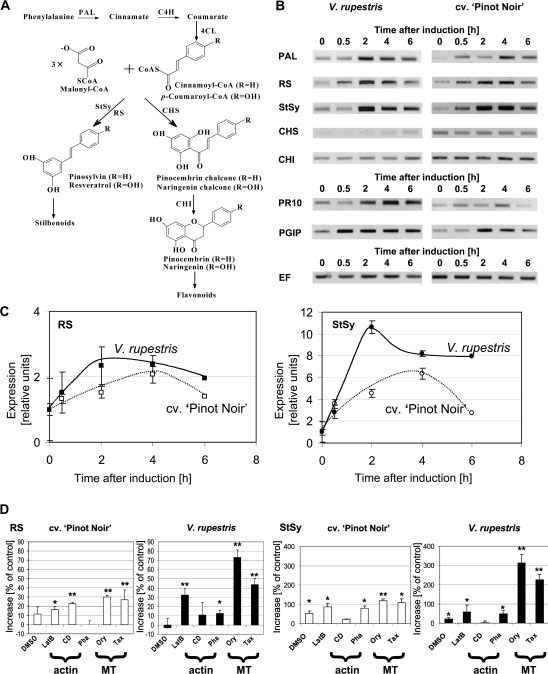
The cytoskeleton undergoes dramatic reorganization during plant defence. This response is generally interpreted as part of the cellular repolarization establishing physical barriers against the invading pathogen. To gain insight into the functional significance of cytoskeletal responses for defence, two Vitis cell cultures that differ in their microtubular dynamics were used, and the cytoskeletal response to the elicitor Harpin in parallel to alkalinization of the medium as a fast response, and the activation of defence-related genes were followed. In one cell line derived from the grapevine cultivar 'Pinot Noir', microtubules contained mostly tyrosinylated alpha-tubulin, indicating high microtubular turnover, whereas in another cell line derived from the wild grapevine V. rupestris, the alpha-tubulin was strongly detyrosinated, indicating low microtubular turnover. The cortical microtubules were disrupted and actin filaments were bundled in both cell lines, but the responses were elevated in V. rupestris as compared with V. vinifera cv. 'Pinot Noir'. The cytoskeletal responsiveness correlated with elicitor-induced alkalinization and the expression of defence genes. Using resveratrol synthase and stilbene synthase as examples, it could be shown that pharmacological manipulation of microtubules could induce gene expression in the absence of elicitor. These findings are discussed with respect to a role for microtubules as positive regulators of defence-induced gene expression.
Figures
References
-
- Abdrakhamanova A, Wang QY, Khokhlova L, Nick P. Is microtubule assembly a trigger for cold acclimation? Plant and Cell Physiology. 2003;44:676–686. - PubMed
-
- Belhadj A, Telef N, Saigne C, Cluzet S, Barrieu F, Hamdi S, Mé rillon JM. Effect of methyl jasmonate in combination with carbohydrates on gene expression of PR proteins, stilbene and anthocyanin accumulation in grapevine cell cultures. Plant Physiology and Biochemistry. 2008;46:493–499. - PubMed
-
- Chuong SDX, Good AG, Taylor GJ, Freeman MC, Moorhead GBG, Muench DG. Large-scale identification of tubulin-binding proteins provides insight on subcellular trafficking, metabolic channeling, and signaling in plant cells. Molecular and Cellular Proteomics. 2004;3:970–983. - PubMed
-
- Coutos-Thévenot P, Poinssot B, Bonomelli A, Yean H, Breda C, Buffard D, Esnault R, Hain R, Boulay M. In vitro tolerance to Botrytis cinerea of grapevine 41B rootstock in transgenic plants expressing the stilbene synthase Vst1 gene under the control of a pathogen-inducible PR 10 promotor. Journal of Experimental Botany. 2001;52:901–910. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
