

A ribosome-associating factor chaperones tail-anchored membrane proteins
- PMID: 20676083
- PMCID: PMC2928861
- DOI: 10.1038/nature09296
A ribosome-associating factor chaperones tail-anchored membrane proteins
Abstract
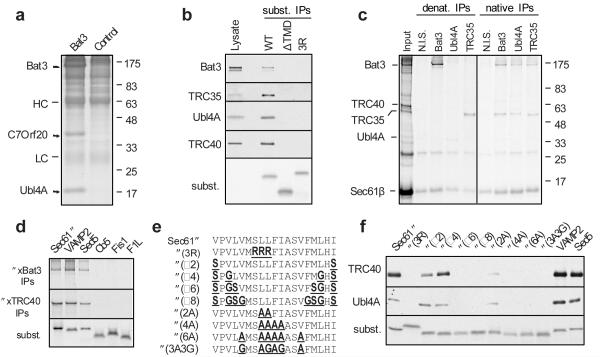
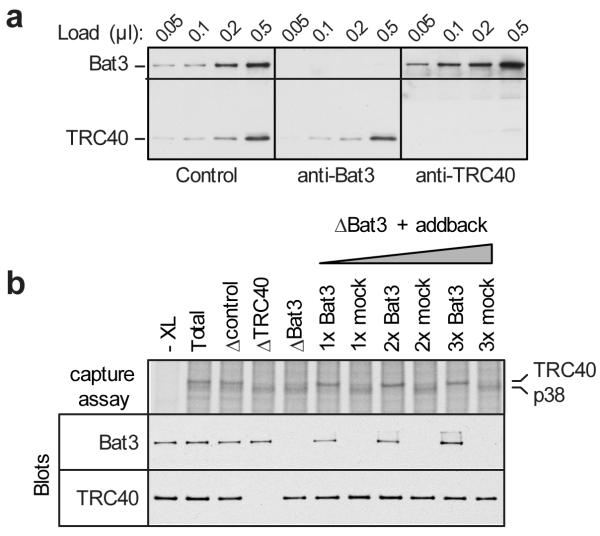
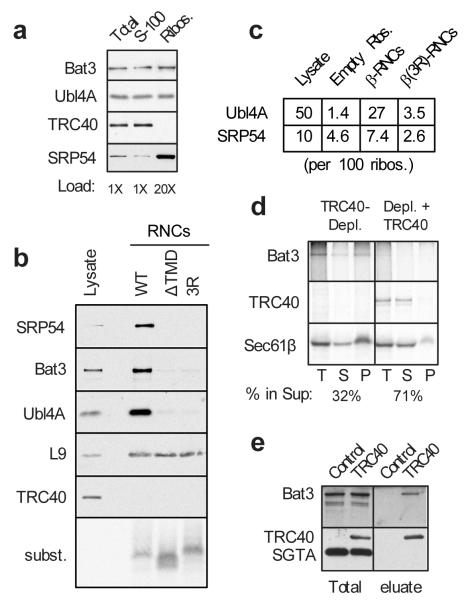
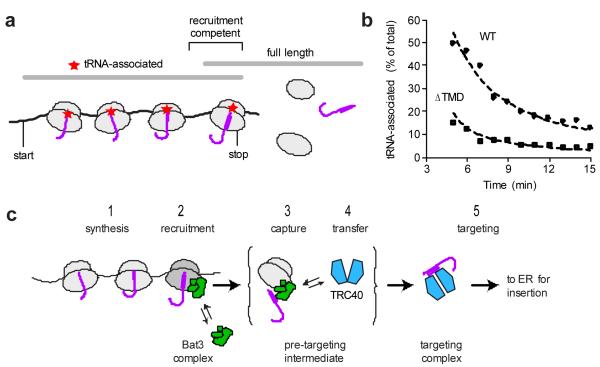
Hundreds of proteins are inserted post-translationally into the endoplasmic reticulum (ER) membrane by a single carboxy-terminal transmembrane domain (TMD). During targeting through the cytosol, the hydrophobic TMD of these tail-anchored (TA) proteins requires constant chaperoning to prevent aggregation or inappropriate interactions. A central component of this targeting system is TRC40, a conserved cytosolic factor that recognizes the TMD of TA proteins and delivers them to the ER for insertion. The mechanism that permits TRC40 to find and capture its TA protein cargos effectively in a highly crowded cytosol is unknown. Here we identify a conserved three-protein complex composed of Bat3, TRC35 and Ubl4A that facilitates TA protein capture by TRC40. This Bat3 complex is recruited to ribosomes synthesizing membrane proteins, interacts with the TMDs of newly released TA proteins, and transfers them to TRC40 for targeting. Depletion of the Bat3 complex allows non-TRC40 factors to compete for TA proteins, explaining their mislocalization in the analogous yeast deletion strains. Thus, the Bat3 complex acts as a TMD-selective chaperone that effectively channels TA proteins to the TRC40 insertion pathway.
Figures
References
-
- Stefanovic S, Hegde RS. Identification of a targeting factor for posttranslational membrane protein insertion into the ER. Cell. 2007;128:1147–59. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
