

Alarm pheromone processing in the ant brain: an evolutionary perspective
- PMID: 20676235
- PMCID: PMC2912167
- DOI: 10.3389/fnbeh.2010.00028
Alarm pheromone processing in the ant brain: an evolutionary perspective
Abstract
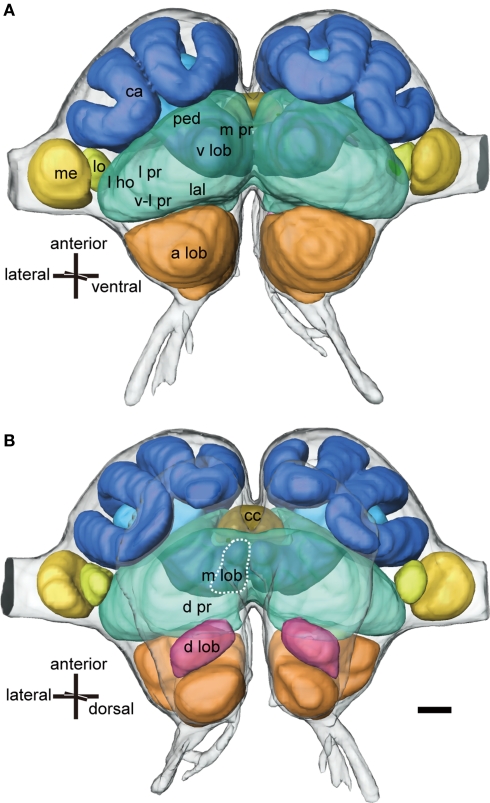
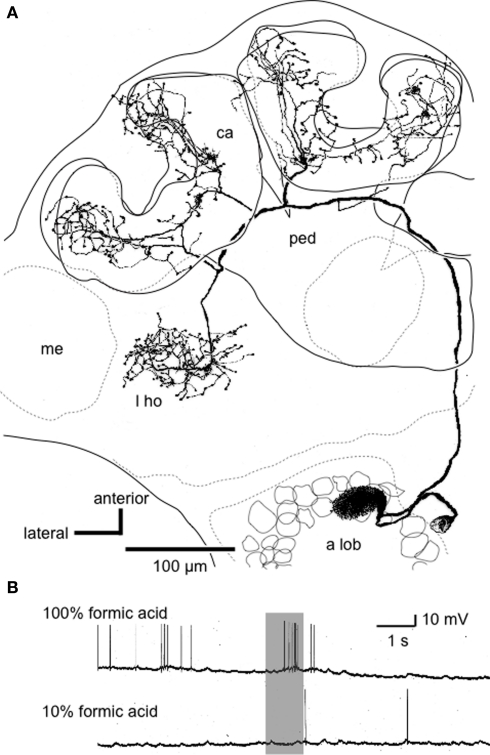
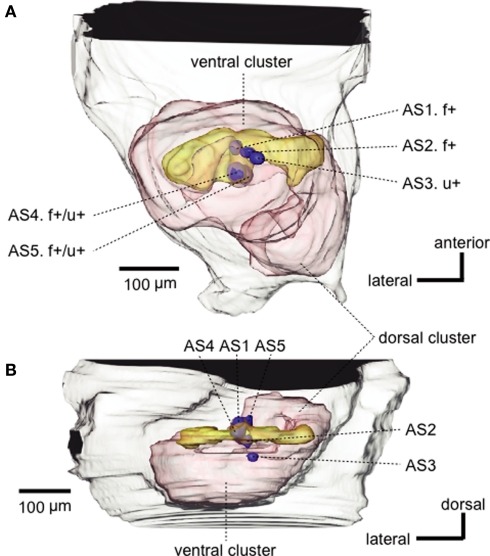
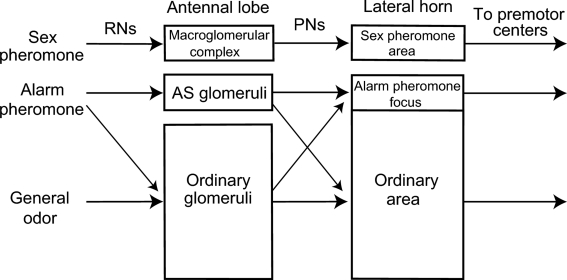
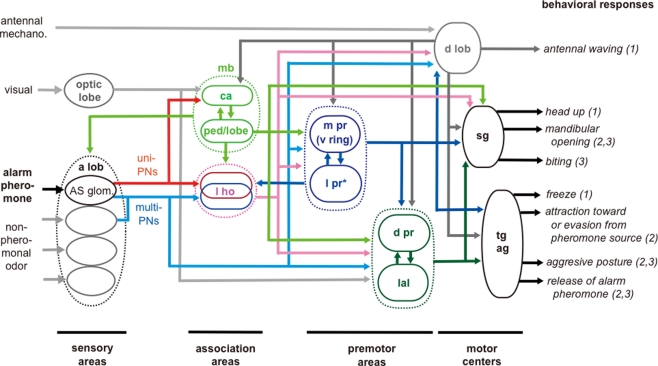
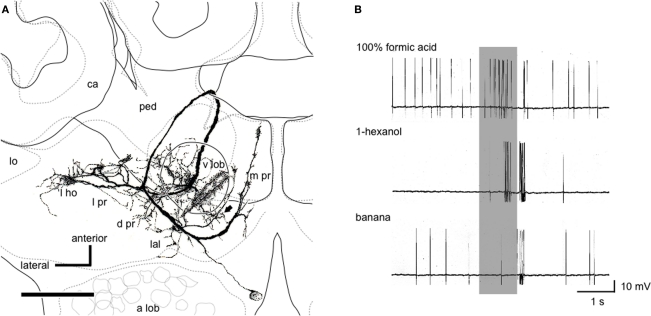
Social insects exhibit sophisticated communication by means of pheromones, one example of which is the use of alarm pheromones to alert nestmates for colony defense. We review recent advances in the understanding of the processing of alarm pheromone information in the ant brain. We found that information about formic acid and n-undecane, alarm pheromone components, is processed in a set of specific glomeruli in the antennal lobe of the ant Camponotus obscuripes. Alarm pheromone information is then transmitted, via projection neurons (PNs), to the lateral horn and the calyces of the mushroom body of the protocerebrum. In the lateral horn, we found a specific area where terminal boutons of alarm pheromone-sensitive PNs are more densely distributed than in the rest of the lateral horn. Some neurons in the protocerebrum responded specifically to formic acid or n-undecane and they may participate in the control of behavioral responses to each pheromone component. Other neurons, especially those originating from the mushroom body lobe, responded also to non-pheromonal odors and may play roles in integration of pheromonal and non-pheromonal signals. We found that a class of neurons receive inputs in the lateral horn and the mushroom body lobe and terminate in a variety of premotor areas. These neurons may participate in the control of aggressive behavior, which is sensitized by alarm pheromones and is triggered by non-pheromonal sensory stimuli associated with a potential enemy. We propose that the alarm pheromone processing system has evolved by differentiation of a part of general odor processing system.
Keywords: aggression; antennal lobe; communication; evolution; mushroom body; pheromone; social insect.
Figures
References
-
- Alaux C., Sinha S., Hasadsri L., Hunt G. J., Guzmán-Novoa E., DeGrandi-Hoffman G., Uribe-Rubio J. L., Southey B. R., Rodriguez-Zas S., Robinson G. E. (2009). Honey bee aggression supports a link between gene regulation and behavioral evolution. Proc. Natl. Acad. Sci. U.S.A. 106, 15400–1540510.1073/pnas.0907043106 - DOI - PMC - PubMed
-
- Billen J. (1994). “Morphology of exocrine glands in social insects: an up-date 100 years after Ch. Janet,” in Les Insectes Sociaux, eds Lenoir A., Arnold A. G., Lepage M. (Paris: Publications Universite Paris Nord; ), p. 214
-
- Blum M. S. (1985). “Alarm pheromone.” in Comparative Insect Physiology, Biochemistry and Pharmacology, Vol 9, eds Kerkut G. A., Gilbert L. I. (Oxford: Pergamon; ) pp. 193–224
-
- Boeckh J., Ernst K. D. (1987). Contribution of single unit analysis in insects to an understanding of olfactory function. J. Comp. Physiol. A. 161, 549–56510.1007/BF00603661 - DOI
LinkOut - more resources
Full Text Sources