

Directional decisions during neutrophil chemotaxis inside bifurcating channels
- PMID: 20676444
- PMCID: PMC3001269
- DOI: 10.1039/c0ib00011f
Directional decisions during neutrophil chemotaxis inside bifurcating channels
Abstract
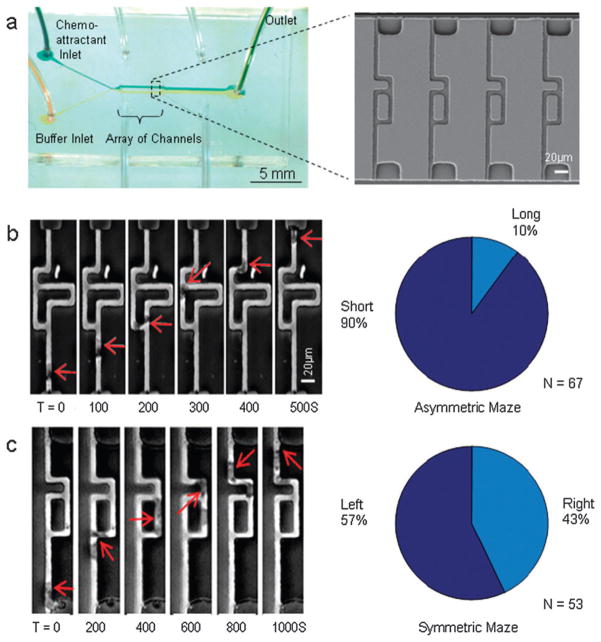
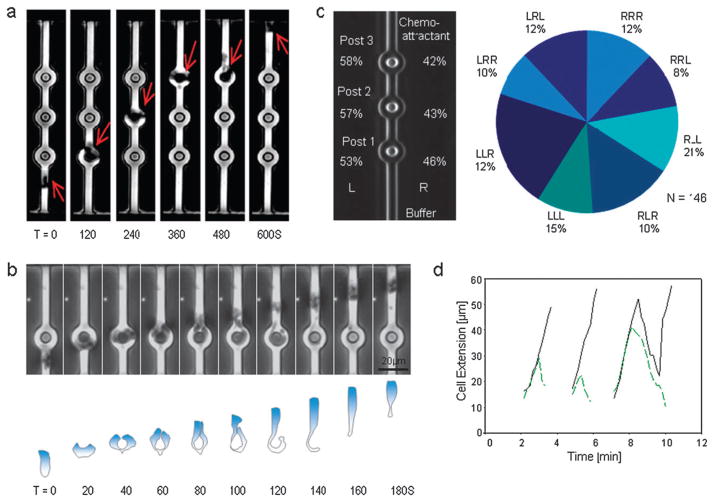
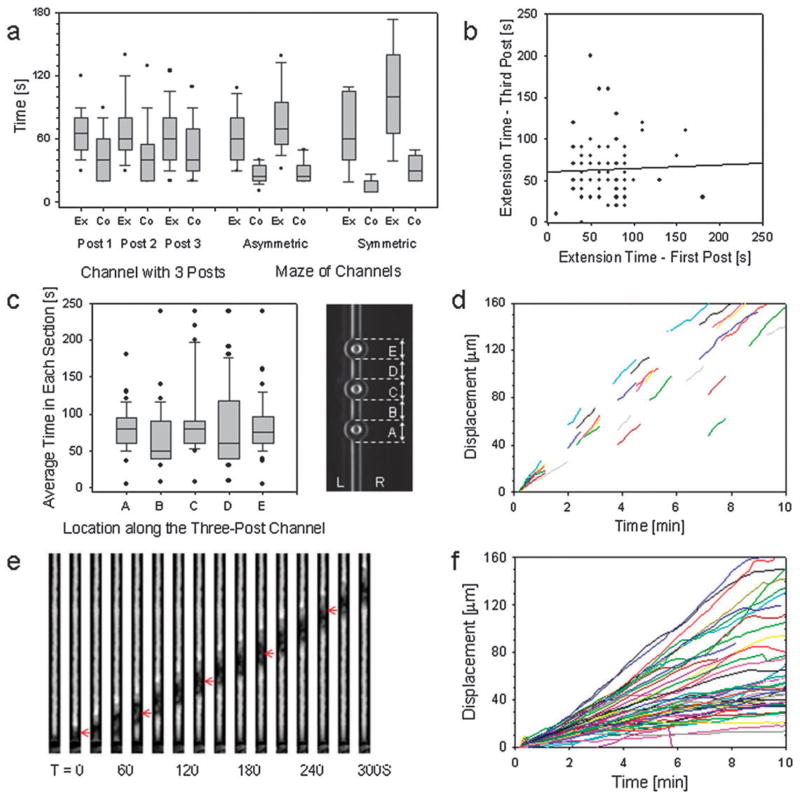
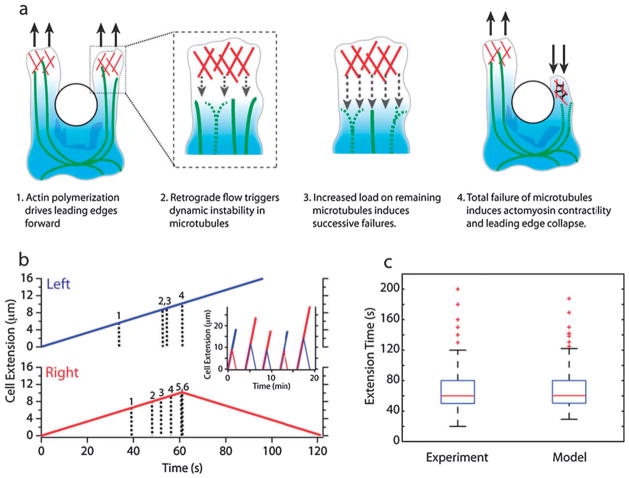
The directional migration of human neutrophils in classical chemotaxis assays is often described as a "biased random walk" implying significant randomness in speed and directionality. However, these experiments are inconsistent with in vivo observations, where neutrophils can navigate effectively through complex tissue microenvironments towards their targets. Here, we demonstrate a novel biomimetic assay for neutrophil chemotaxis using enclosed microfluidic channels. Remarkably, under these enclosed conditions, neutrophils recapitulate the highly robust and efficient navigation observed in vivo. In straight channels, neutrophils undergo sustained, unidirectional motion towards a chemoattractant source. In more complex maze-like geometries, neutrophils are able to select the most direct route over 90% of the time. Finally, at symmetric bifurcations, neutrophils split their leading edge into two sections and a "tug of war" ensues. The competition between the two new leading edges is ultimately resolved by stochastic, symmetry-breaking behavior. This behavior is suggestive of directional decision-making localized at the leading edge and a signaling role played by the cellular cytoskeleton.
Figures
References
-
- Bodey GP, Buckley M, Sathe YS, Freireich EJ. Ann Intern Med. 1966;64:328–340. - PubMed
-
- Tharp WG, Yadav R, Irimia D, Upadhyaya A, Samadani A, Hurtado O, Liu SY, Munisamy S, Brainard DM, Mahon MJ, Nourshargh S, van Oudenaarden A, Toner MG, Poznansky MC. J Leukocyte Biol. 2006;79:539–554. - PubMed
-
- Mathias JR, Walters KB, Huttenlocher A. Methods Mol Biol. 2009;571:151–166. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
