

Identification of a small molecule inhibitor of importin β mediated nuclear import by confocal on-bead screening of tagged one-bead one-compound libraries
- PMID: 20677820
- PMCID: PMC2956136
- DOI: 10.1021/cb100094k
Identification of a small molecule inhibitor of importin β mediated nuclear import by confocal on-bead screening of tagged one-bead one-compound libraries
Abstract
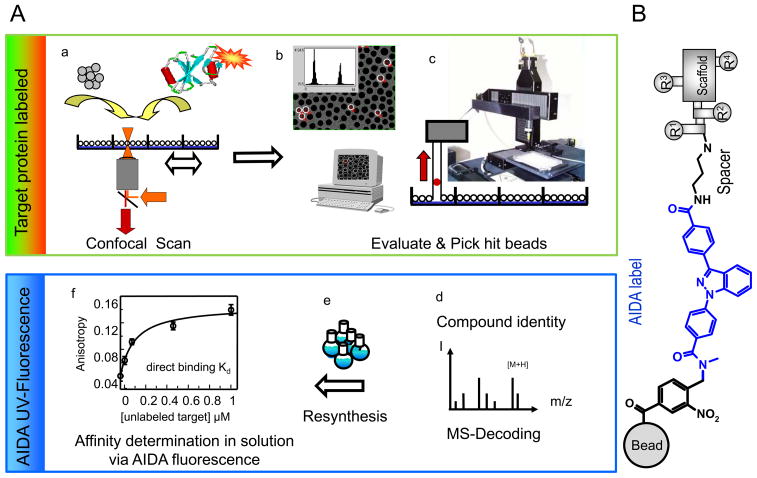
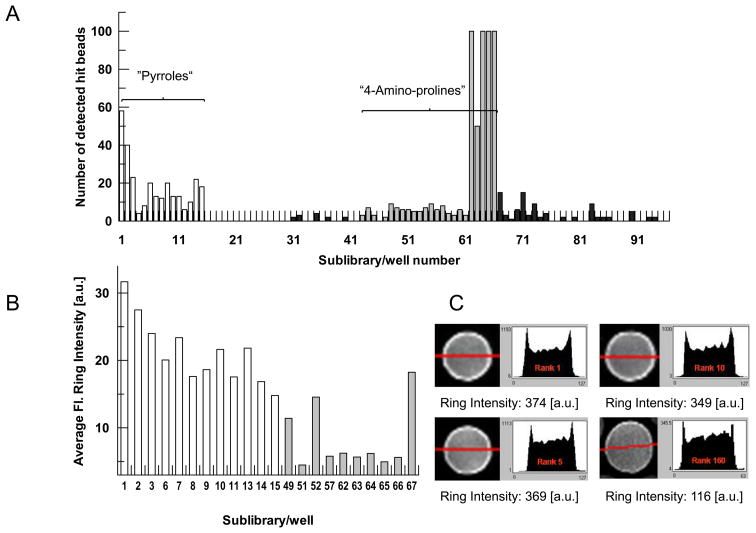
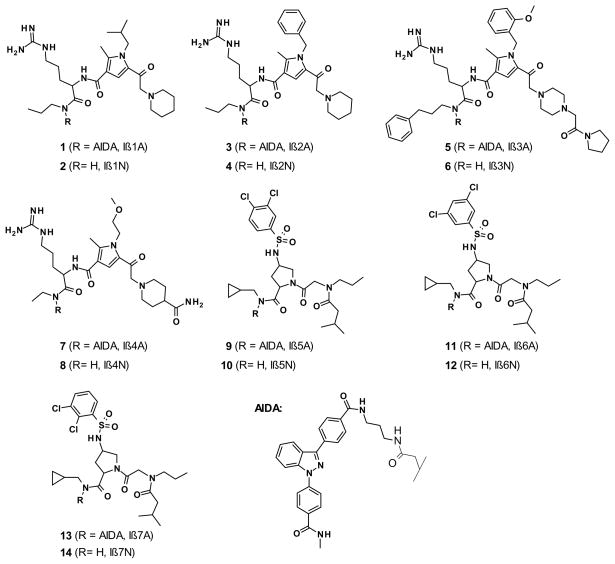
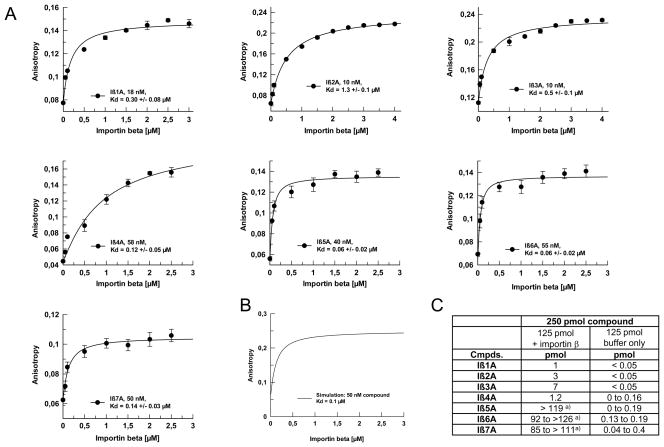
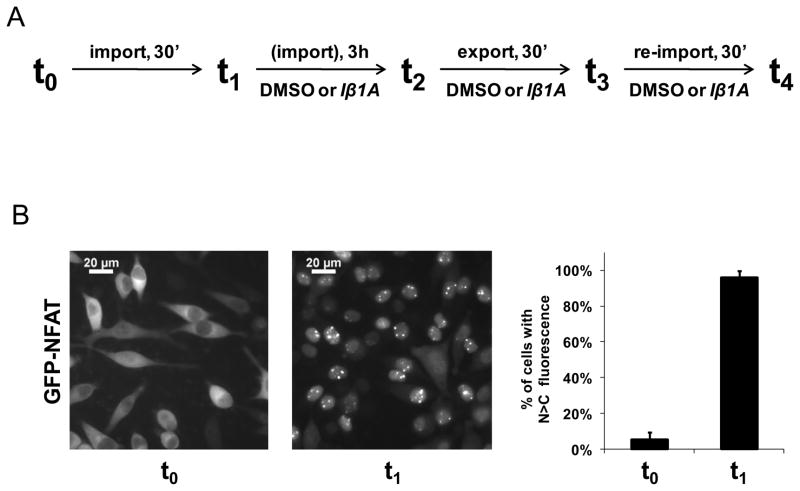
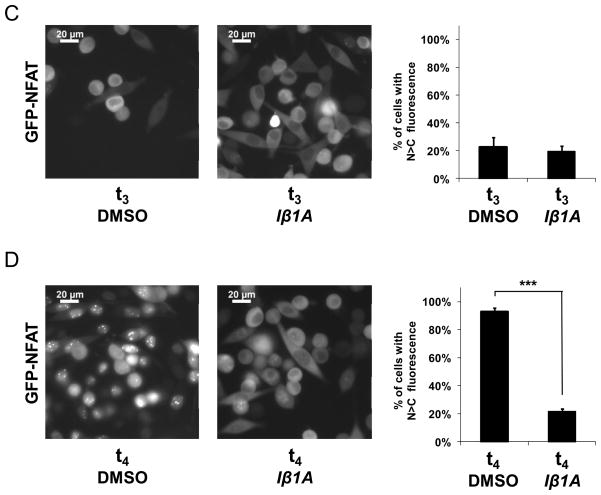
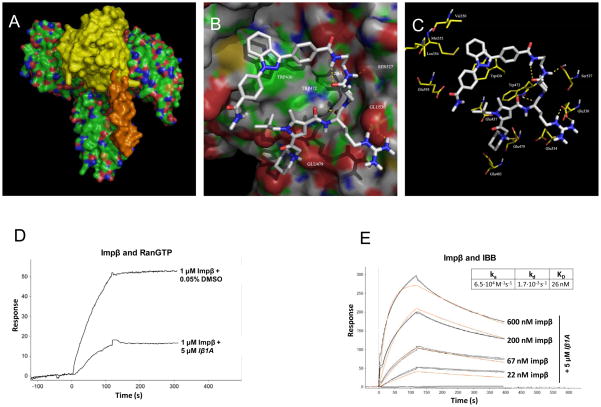
In eukaryotic cells, proteins and RNAs are transported between the nucleus and the cytoplasm by nuclear import and export receptors. Over the past decade, small molecules that inhibit the nuclear export receptor CRM1 have been identified, most notably leptomycin B. However, up to now no small molecule inhibitors of nuclear import have been described. Here we have used our automated confocal nanoscanning and bead picking method (CONA) for on-bead screening of a one-bead one-compound library to identify the first such import inhibitor, karyostatin 1A. Karyostatin 1A binds importin β with high nanomolar affinity and specifically inhibits importin α/β mediated nuclear import at low micromolar concentrations in vitro and in living cells, without perturbing transportin mediated nuclear import or CRM1 mediated nuclear export. Surface plasmon resonance binding experiments suggest that karyostatin 1A acts by disrupting the interaction between importin β and the GTPase Ran. As a selective inhibitor of the importin α/β import pathway, karyostatin 1A will provide a valuable tool for future studies of nucleocytoplasmic trafficking.
Figures

References
-
- Terry LJ, Shows EB, Wente SR. Crossing the nuclear envelope: hierarchical regulation of nucleocytoplasmic transport. Science. 2007;318:1412–1416. - PubMed
-
- Stewart M. Molecular mechanism of the nuclear protein import cycle. Nat Rev Mol Cell Biol. 2007;8:195–208. - PubMed
-
- Weis K. Importins and exportins: how to get in and out of the nucleus. Trends Biochem Sci. 1998;23:235. - PubMed
-
- Cook A, Bono F, Jinek M, Conti E. Structural biology of nucleocytoplasmic transport. Annu Rev Biochem. 2007;76:647–671. - PubMed
-
- Bednenko J, Cingolani G, Gerace L. Nucleocytoplasmic transport: navigating the channel. Traffic. 2003;4:127–135. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
