

Toxoplasma gondii protease TgSUB1 is required for cell surface processing of micronemal adhesive complexes and efficient adhesion of tachyzoites
- PMID: 20678172
- PMCID: PMC2997387
- DOI: 10.1111/j.1462-5822.2010.01509.x
Toxoplasma gondii protease TgSUB1 is required for cell surface processing of micronemal adhesive complexes and efficient adhesion of tachyzoites
Abstract
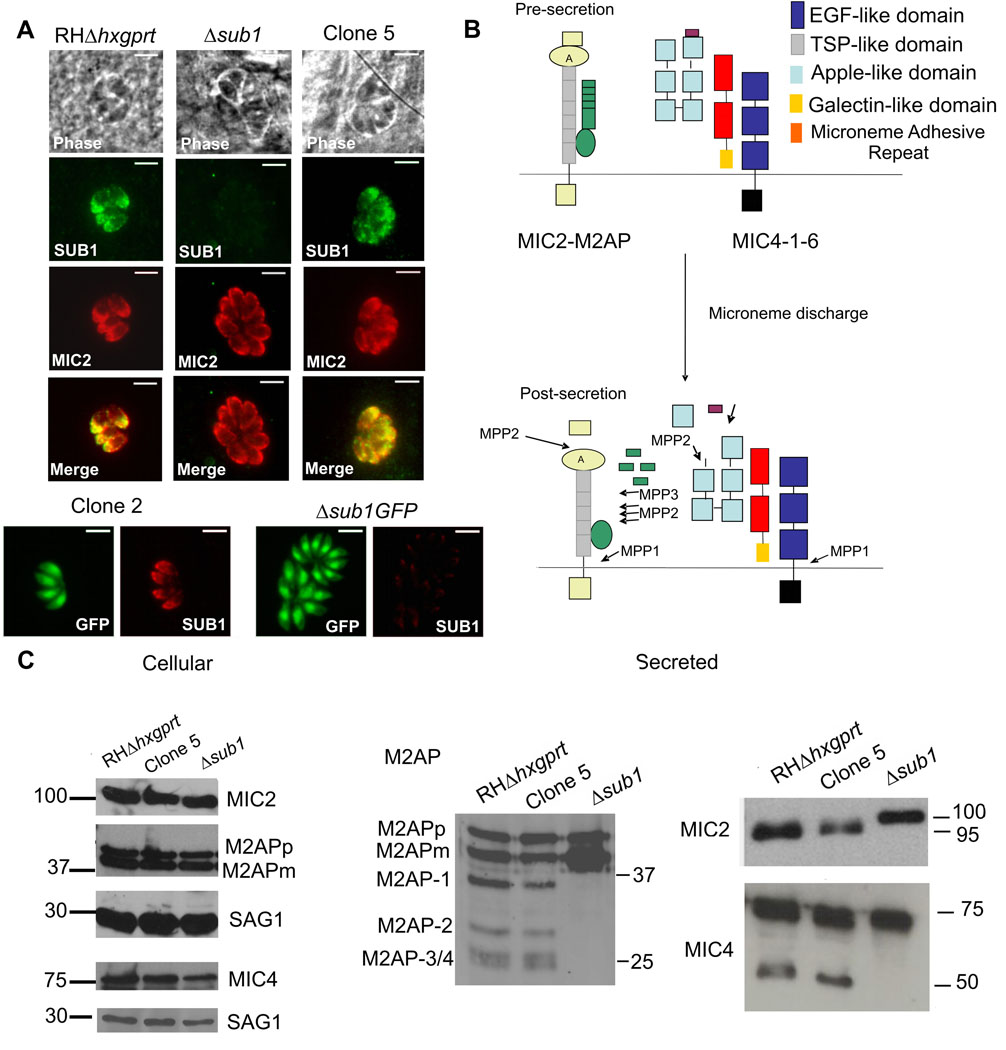
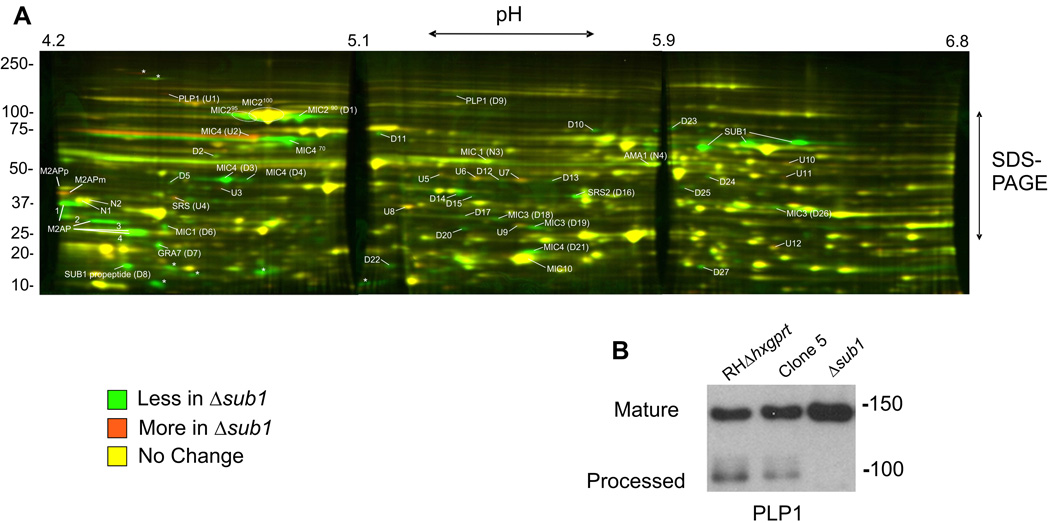
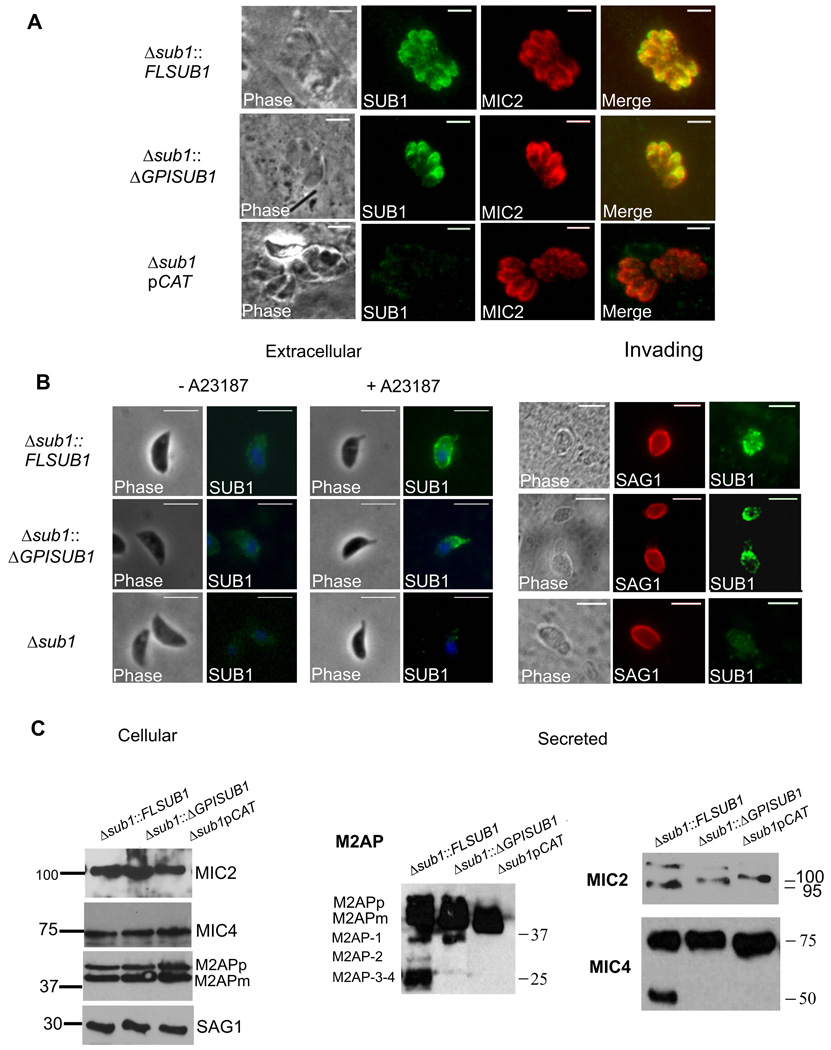
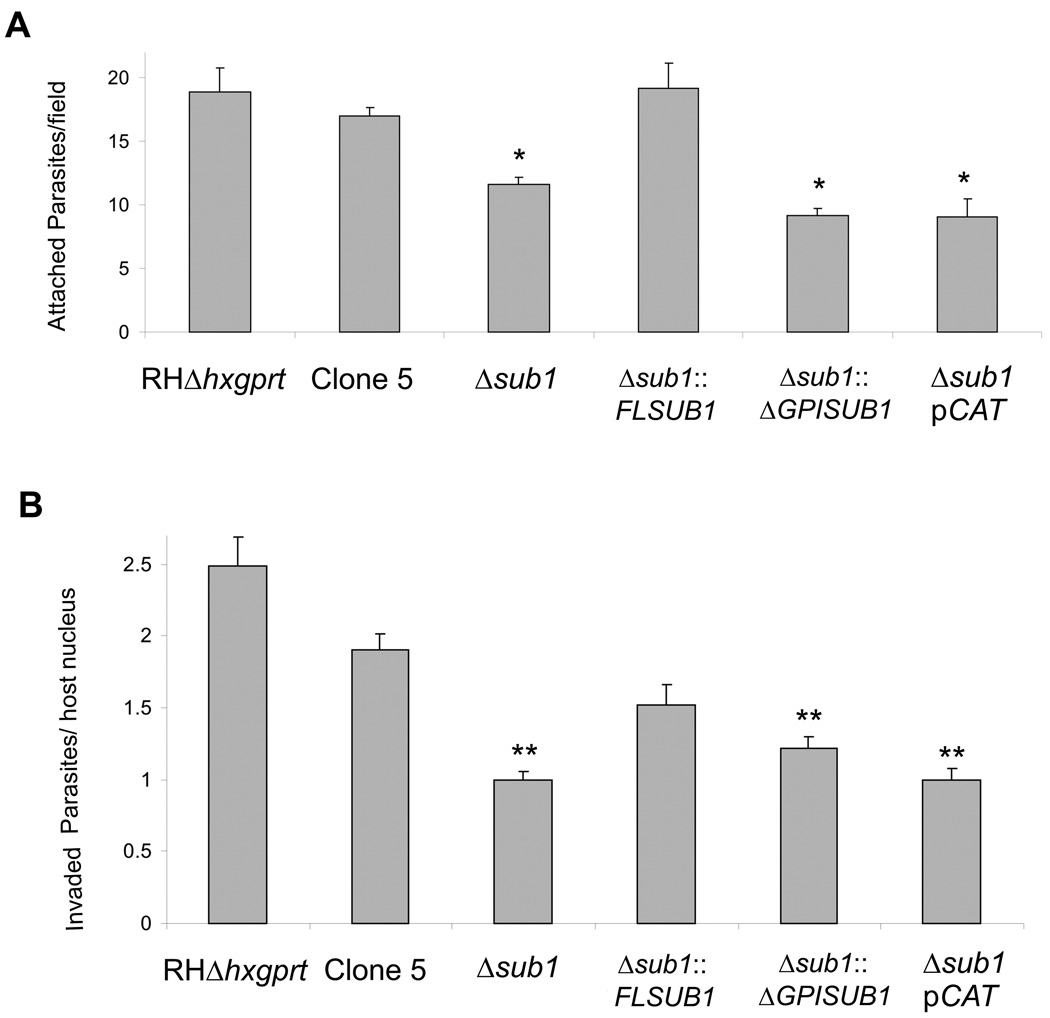
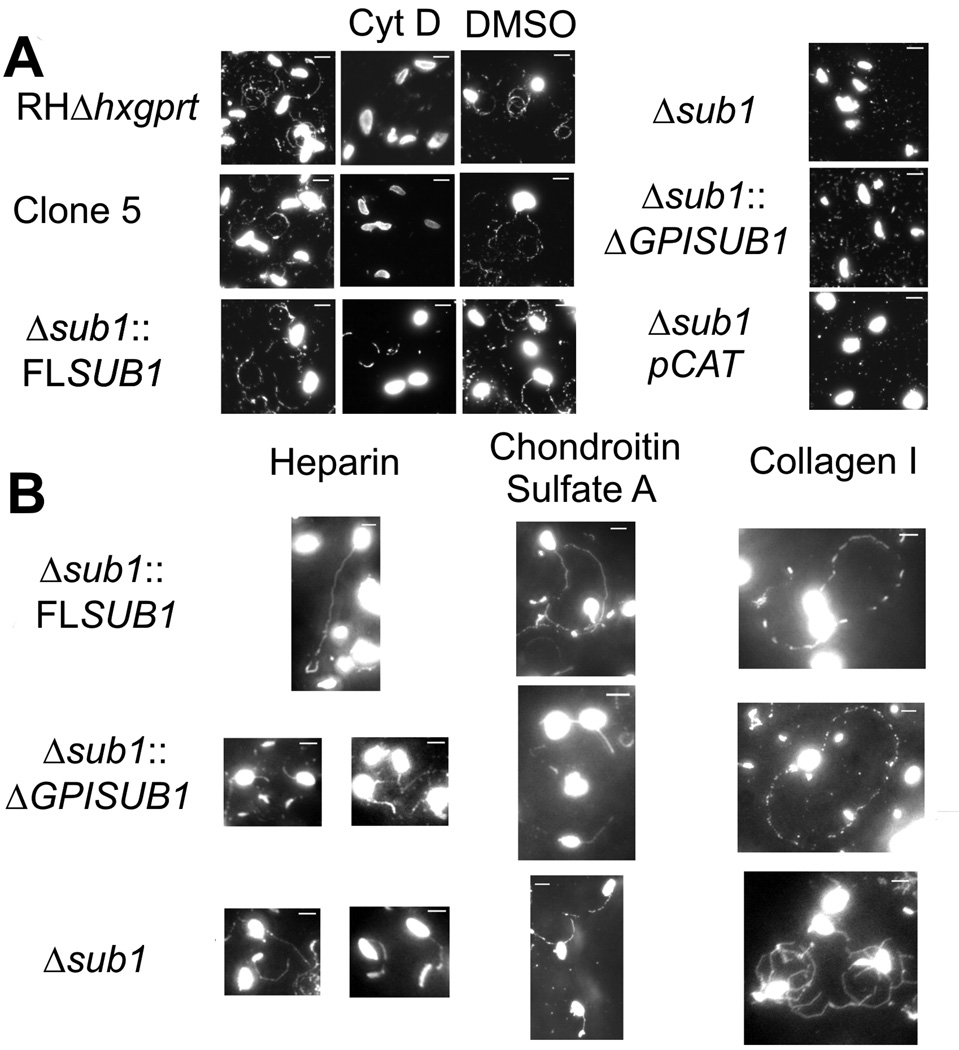
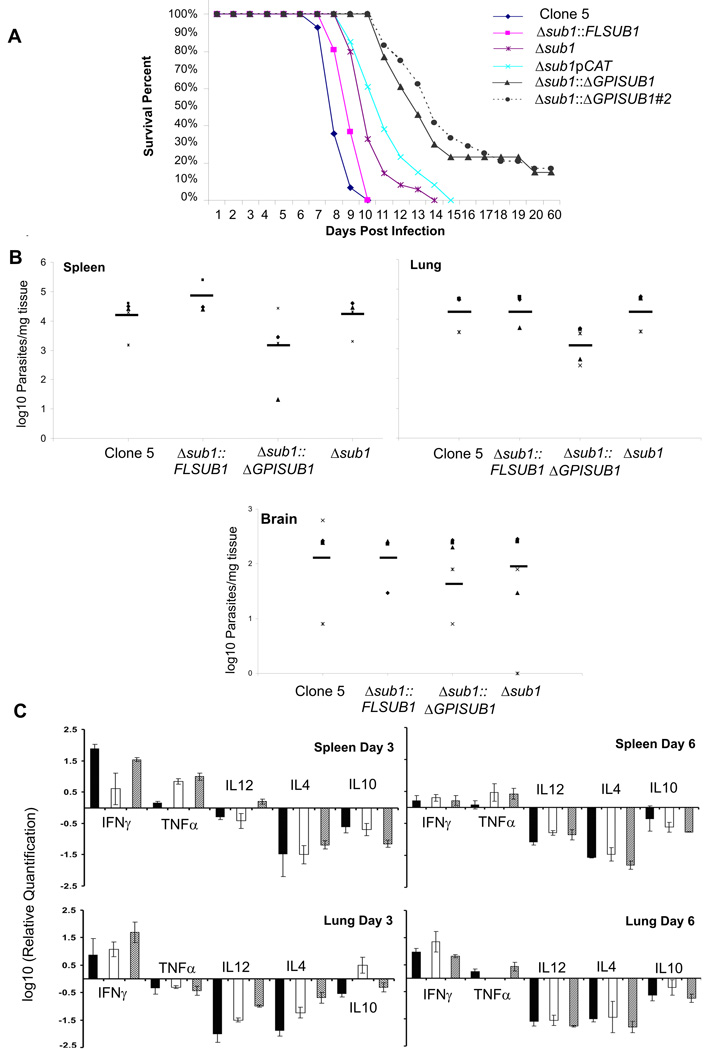
Host cell invasion by Toxoplasma gondii is critically dependent upon adhesive proteins secreted from the micronemes. Proteolytic trimming of microneme contents occurs rapidly after their secretion onto the parasite surface and is proposed to regulate adhesive complex activation to enhance binding to host cell receptors. However, the proteases responsible and their exact function are still unknown. In this report, we show that T. gondii tachyzoites lacking the microneme subtilisin protease TgSUB1 have a profound defect in surface processing of secreted microneme proteins. Notably parasites lack protease activity responsible for proteolytic trimming of MIC2, MIC4 and M2AP after release onto the parasite surface. Although complementation with full-length TgSUB1 restores processing, complementation of Δsub1 parasites with TgSUB1 lacking the GPI anchor (Δsub1::ΔGPISUB1) only partially restores microneme protein processing. Loss of TgSUB1 decreases cell attachment and in vitro gliding efficiency leading to lower initial rates of invasion. Δsub1 and Δsub1::ΔGPISUB1 parasites are also less virulent in mice. Thus TgSUB1 is involved in micronemal protein processing and regulation of adhesive properties of macromolecular adhesive complexes involved in host cell invasion.
© 2010 Blackwell Publishing Ltd.
Figures
Similar articles
-
The prodomain of Toxoplasma gondii GPI-anchored subtilase TgSUB1 mediates its targeting to micronemes.Traffic. 2008 Sep;9(9):1485-96. doi: 10.1111/j.1600-0854.2008.00774.x. Epub 2008 Jun 2. Traffic. 2008. PMID: 18532988 Free PMC article.
-
A cathepsin C-like protease mediates the post-translation modification of Toxoplasma gondii secretory proteins for optimal invasion and egress.mBio. 2023 Aug 31;14(4):e0017423. doi: 10.1128/mbio.00174-23. Epub 2023 Jun 16. mBio. 2023. PMID: 37326431 Free PMC article.
-
Preparing for an invasion: charting the pathway of adhesion proteins to Toxoplasma micronemes.Parasitol Res. 2006 Apr;98(5):389-95. doi: 10.1007/s00436-005-0062-2. Epub 2005 Dec 30. Parasitol Res. 2006. PMID: 16385407 Review.
-
Functional analysis of rhomboid proteases during Toxoplasma invasion.mBio. 2014 Oct 21;5(5):e01795-14. doi: 10.1128/mBio.01795-14. mBio. 2014. PMID: 25336455 Free PMC article.
-
Microneme proteins: structural and functional requirements to promote adhesion and invasion by the apicomplexan parasite Toxoplasma gondii.Int J Parasitol. 2001 Oct;31(12):1293-302. doi: 10.1016/s0020-7519(01)00257-0. Int J Parasitol. 2001. PMID: 11566297 Review.
Cited by
-
Differential gene expression in mice infected with distinct Toxoplasma strains.Infect Immun. 2012 Mar;80(3):968-74. doi: 10.1128/IAI.05421-11. Epub 2011 Dec 5. Infect Immun. 2012. PMID: 22144491 Free PMC article.
-
The Rhoptry Pseudokinase ROP54 Modulates Toxoplasma gondii Virulence and Host GBP2 Loading.mSphere. 2016 Mar 23;1(2):e00045-16. doi: 10.1128/mSphere.00045-16. eCollection 2016 Mar-Apr. mSphere. 2016. PMID: 27303719 Free PMC article.
-
Factors mediating plastid dependency and the origins of parasitism in apicomplexans and their close relatives.Proc Natl Acad Sci U S A. 2015 Aug 18;112(33):10200-7. doi: 10.1073/pnas.1423790112. Epub 2015 Feb 25. Proc Natl Acad Sci U S A. 2015. PMID: 25717057 Free PMC article.
-
Prodomain-driven enzyme dimerization: a pH-dependent autoinhibition mechanism that controls Plasmodium Sub1 activity before merozoite egress.mBio. 2024 Mar 13;15(3):e0019824. doi: 10.1128/mbio.00198-24. Epub 2024 Feb 22. mBio. 2024. PMID: 38386597 Free PMC article.
-
Transcriptional profiling of Toll-like receptor 2-deficient primary murine brain cells during Toxoplasma gondii infection.PLoS One. 2017 Nov 14;12(11):e0187703. doi: 10.1371/journal.pone.0187703. eCollection 2017. PLoS One. 2017. PMID: 29136637 Free PMC article.
References
-
- Barragan A, Brossier F, Sibley DL. Transepithelial migration of toxoplasma gondii involves an interaction of intercellular adhesion molecule 1 (ICAM-1) with the parasite adhesin MIC2. Cell. Microbiol. 2005;7:561–568. - PubMed
-
- Baum J, Papenfuss A, Baum B, Speed T, Cowman A. Regulation of apicomplexan actin-based motility. Nat. Rev. Microbiol. 2006;4:621–628. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
