

Regulated protein kinases and phosphatases in cell cycle decisions
- PMID: 20678910
- PMCID: PMC3769698
- DOI: 10.1016/j.ceb.2010.07.001
Regulated protein kinases and phosphatases in cell cycle decisions
Abstract
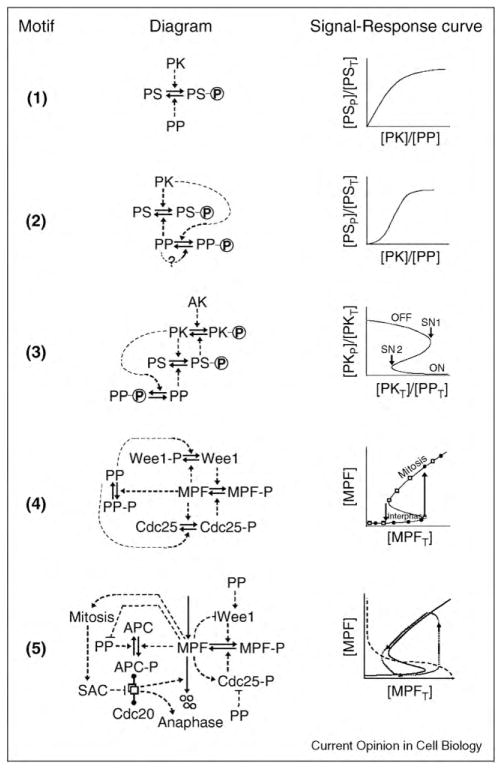
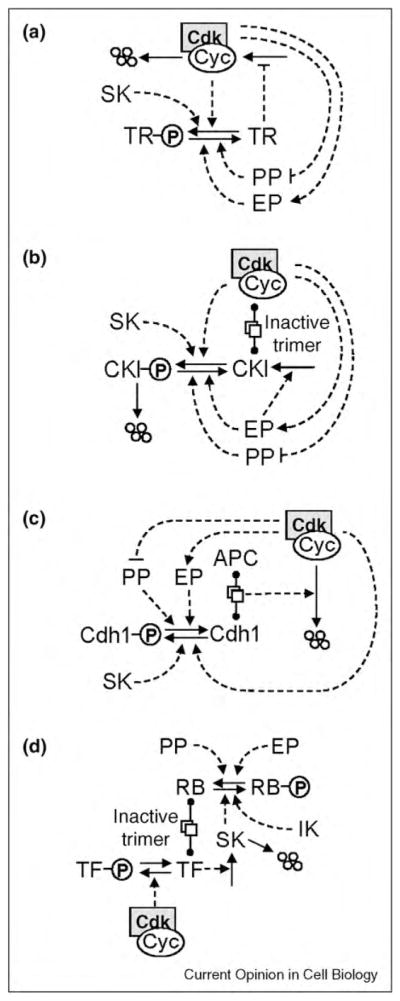
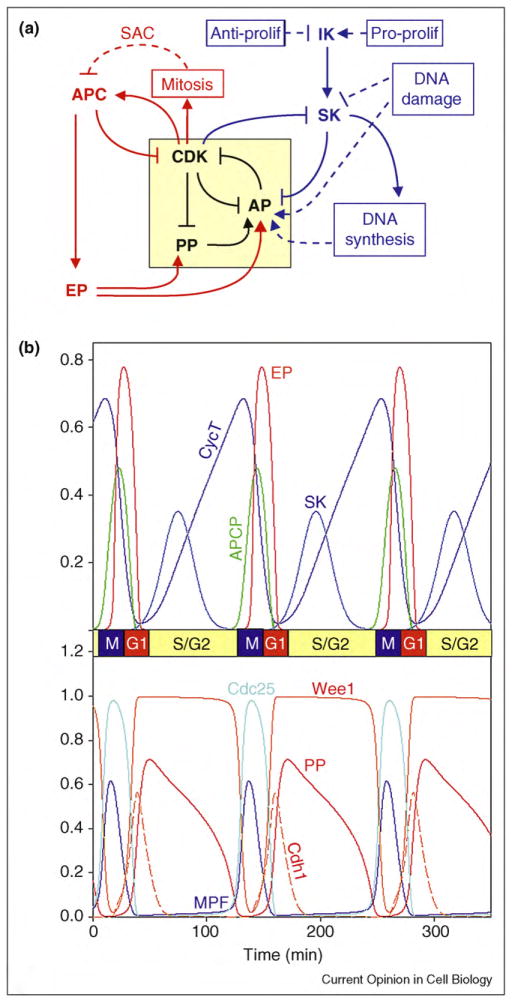
Many aspects of cell physiology are controlled by protein kinases and phosphatases, which together determine the phosphorylation state of targeted substrates. Some of these target proteins are themselves kinases or phosphatases or other components of a regulatory network characterized by feedback and feed-forward loops. In this review we describe some common regulatory motifs involving kinases, phosphatases, and their substrates, focusing particularly on bistable switches involved in cellular decision processes. These general principles are applied to cell cycle transitions, with special emphasis on the roles of regulated phosphatases in orchestrating progression from one phase to the next of the DNA replication-division cycle.
Copyright © 2010 Elsevier Ltd. All rights reserved.
Figures
References
-
- Bray D. Protein molecules as computational elements in living cells. Nature. 1995;376:307–312. - PubMed
-
- Tyson JJ, Chen K, Novak B. Network dynamics and cell physiology. Nat Rev Mol Cell Biol. 2001;2:908–916. - PubMed
-
- Bollen M, Gerlich DW, Lesage B. Mitotic phosphatases: from entry guards to exit guides. Trends Cell Biol. 2009;19:531–541. - PubMed
-
- De Wulf P, Montani F, Visintin R. Protein phosphatases take the mitotic stage. Curr Opin Cell Biol. 2009;21:806–815. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
