

Different functions of the C3HC4 zinc RING finger peroxins PEX10, PEX2, and PEX12 in peroxisome formation and matrix protein import
- PMID: 20679226
- PMCID: PMC2930433
- DOI: 10.1073/pnas.1009174107
Different functions of the C3HC4 zinc RING finger peroxins PEX10, PEX2, and PEX12 in peroxisome formation and matrix protein import
Abstract
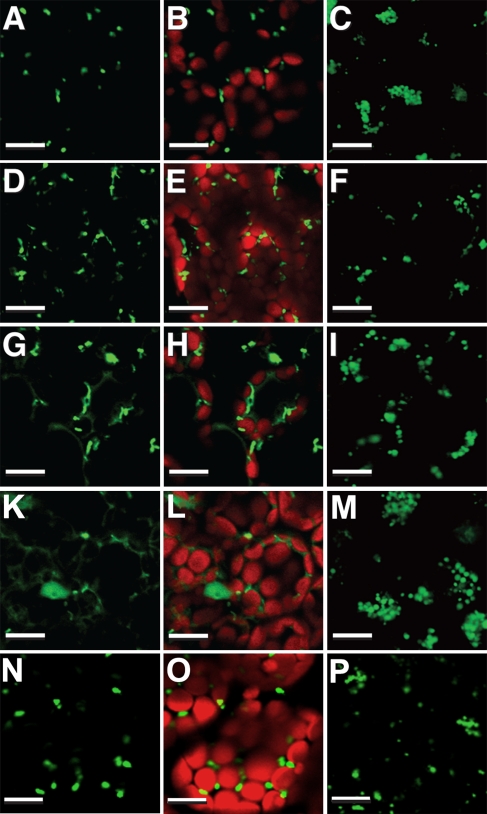
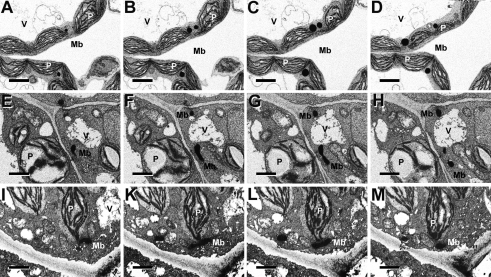
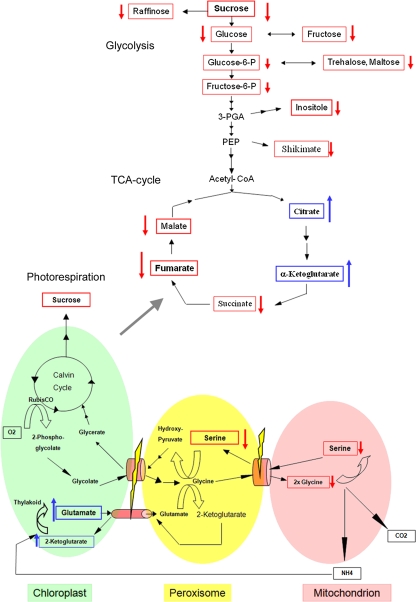
The integral peroxisomal membrane proteins PEX10, PEX2, and PEX12 contain a zinc RING finger close to the C terminus. Loss of function of these peroxins causes embryo lethality at the heart stage in Arabidopsis. Preventing the coordination of Zn(2+) ions by amino acid substitutions in PEX10, PEX2, and PEX12 and overexpressing the resulting conditional sublethal mutations in WT uncovered additional functions of PEX10. Plants overexpressing DeltaZn-mutant PEX10 display deformed peroxisomal shapes causing diminished contact with chloroplasts and possibly with mitochondria. These changes correlated with impaired metabolite transfer and, at high CO(2), recoverable defective photorespiration plus dwarfish phenotype. The N-terminal PEX10 domain is critical for peroxisome biogenesis and plant development. A point mutation in the highly conserved TLGEEY motif results in vermiform peroxisome shape without impairing organelle contact. Addition of an N-terminal T7 tag to WT PEX0 resulted in partially recoverable reduced growth and defective inflorescences persisting under high CO(2). In contrast, plants overexpressing PEX2-DeltaZn-T7 grow like WT in normal atmosphere, contain normal-shaped peroxisomes, but display impaired peroxisomal matrix protein import. PEX12-DeltaZn-T7 mutants exhibit unimpaired import of matrix protein and normal-shaped peroxisomes when grown in normal atmosphere. During seed germination, glyoxysomes form a reticulum around the lipid bodies for mobilization of storage oil. The formation of this glyoxysomal reticulum seemed to be impaired in PEX10-DeltaZn but not in PEX2-DeltaZn-T7 or PEX12-DeltaZn-T7 plants. Both cytosolic PEX10 domains seem essential for peroxisome structure but differ in metabolic function, suggesting a role for this plant peroxin in addition to the import of matrix protein via ubiquitination of PEX5.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

Similar articles
-
Requirement of the C3HC4 zinc RING finger of the Arabidopsis PEX10 for photorespiration and leaf peroxisome contact with chloroplasts.Proc Natl Acad Sci U S A. 2007 Jan 16;104(3):1069-74. doi: 10.1073/pnas.0610402104. Epub 2007 Jan 10. Proc Natl Acad Sci U S A. 2007. PMID: 17215364 Free PMC article.
-
Peroxisomal ubiquitin-protein ligases peroxin2 and peroxin10 have distinct but synergistic roles in matrix protein import and peroxin5 retrotranslocation in Arabidopsis.Plant Physiol. 2014 Nov;166(3):1329-44. doi: 10.1104/pp.114.247148. Epub 2014 Sep 11. Plant Physiol. 2014. PMID: 25214533 Free PMC article.
-
Genetic Interactions between PEROXIN12 and Other Peroxisome-Associated Ubiquitination Components.Plant Physiol. 2016 Nov;172(3):1643-1656. doi: 10.1104/pp.16.01211. Epub 2016 Sep 20. Plant Physiol. 2016. PMID: 27650450 Free PMC article.
-
Peroxisome biogenesis, protein targeting mechanisms and PEX gene functions in plants.Biochim Biophys Acta. 2016 May;1863(5):850-62. doi: 10.1016/j.bbamcr.2015.09.027. Epub 2015 Sep 25. Biochim Biophys Acta. 2016. PMID: 26408938 Review.
-
Biochemical and molecular approaches to understanding protein import into peroxisomes.Biochem Soc Trans. 2000;28(4):499-504. Biochem Soc Trans. 2000. PMID: 10961948 Review.
Cited by
-
Production of omega-3 eicosapentaenoic acid by metabolic engineering of Yarrowia lipolytica.Nat Biotechnol. 2013 Aug;31(8):734-40. doi: 10.1038/nbt.2622. Epub 2013 Jul 21. Nat Biotechnol. 2013. PMID: 23873085
-
Defining the plant peroxisomal proteome: from Arabidopsis to rice.Front Plant Sci. 2011 Dec 27;2:103. doi: 10.3389/fpls.2011.00103. eCollection 2011. Front Plant Sci. 2011. PMID: 22645559 Free PMC article.
-
Peroxisome Function, Biogenesis, and Dynamics in Plants.Plant Physiol. 2018 Jan;176(1):162-177. doi: 10.1104/pp.17.01050. Epub 2017 Oct 11. Plant Physiol. 2018. PMID: 29021223 Free PMC article. Review.
-
Genome-wide identification of RING finger genes in flax (Linum usitatissimum) and analyses of their evolution.PeerJ. 2021 Nov 15;9:e12491. doi: 10.7717/peerj.12491. eCollection 2021. PeerJ. 2021. PMID: 34820204 Free PMC article.
-
FgPEX1 and FgPEX10 are required for the maintenance of Woronin bodies and full virulence of Fusarium graminearum.Curr Genet. 2019 Dec;65(6):1383-1396. doi: 10.1007/s00294-019-00994-8. Epub 2019 May 20. Curr Genet. 2019. PMID: 31111312
References
-
- Brown LA, Baker A. Shuttles and cycles: Transport of proteins into the peroxisome matrix (review) Mol Membr Biol. 2008;25:363–375. - PubMed
-
- Reumann S, Weber AP. Plant peroxisomes respire in the light: Some gaps of the photorespiratory C2 cycle have become filled—others remain. Biochim Biophys Acta. 2006;1763:1496–1510. - PubMed
-
- Hayashi M, Nishimura M. Arabidopsis thaliana—a model organism to study plant peroxisomes. Biochim Biophys Acta. 2006;1763:1382–1391. - PubMed
-
- Mano S, Nishimura M. Plant peroxisomes. Vitam Horm. 2005;72:111–154. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
