

Review
doi: 10.1101/cshperspect.a002188.
Epub 2010 Apr 21.
Membrane transport in primitive cells
Affiliations
- PMID: 20679338
- PMCID: PMC2908771
- DOI: 10.1101/cshperspect.a002188
Item in Clipboard
Review
Membrane transport in primitive cells
Cold Spring Harb Perspect Biol.
2010 Aug.
Abstract
Although model protocellular membranes consisting of monoacyl lipids are similar to membranes composed of contemporary diacyl lipids, they differ in at least one important aspect. Model protocellular membranes allow for the passage of polar solutes and thus can potentially support cell-to functions without the aid of transport machinery. The ability to transport polar molecules likely stems from increased lipid dynamics. Selectively permeable vesicle membranes composed of monoacyl lipids allow for many lifelike processes to emerge from a remarkably small set of molecules.
Figures
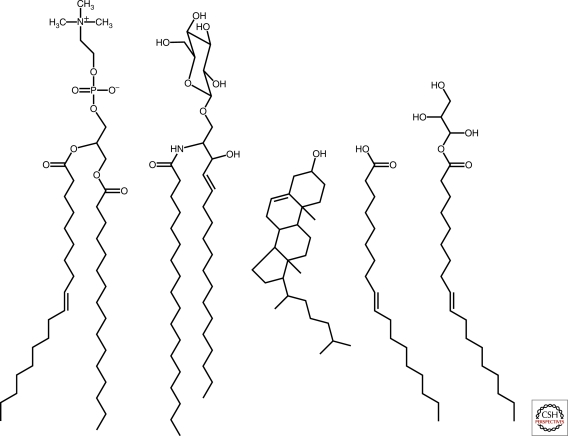
Lipid chemical structures. From left to right, phospholipid (1-palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine), sphingolipid (glucosyl ceramide), sterol (cholesterol), fatty acid (oleate), and glycerol monoester of fatty acid (monoolein). Phospholipids, spingolipids, and sterols are common components of biological membranes, whereas fatty acids and glycerol monoesters of fatty acids are constituents of model protocellular membranes.
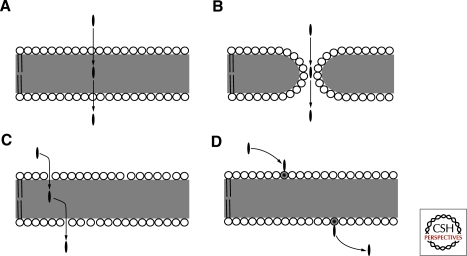
Membrane transport mechanisms. (A) Solubility-diffusion model. (B) Transient pore model. (C) Head-group gated model. (D) Lipid flipping-carrier model. Circles represent lipid head groups and the gray area indicates the hydrophobic region occupied by lipid acyl chains. The black oval is the solute molecule.
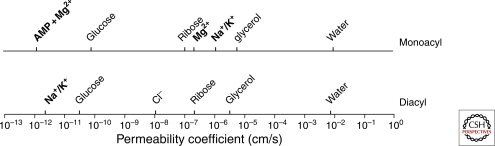
Comparison between solute permeability coefficients of liquid-crystalline diacyl phospholipid and monoacyl fatty acid membranes. Solutes in bold type permeate differently through both membranes. AMP permeation depends on the presence of Mg++. AMP and Mg++ permeability through diacyl phospholipid membranes does not occur within the time scale represented by the x-axis. Differences in permeability between monoacyl and diacyl lipid membranes are small for nonpolar solutes and large for polar solutes. (Data were compiled from Paula et al. 1996; Paula et al. 1998; Paula and Deamer 1999; Chen and Szostak 2004b; Chen et al. 2005; Sacerdote and Szostak 2005; Mansy et al. 2008.)
References
-
- Armstrong VT, Brzustowicz MR, Wassall SR, Jenski LJ, Stillwell W 2003. Rapid flip-flop in polyunsaturated (docosahexaenoate) phospholipid membranes. Arch Biochem Biophys 414: 74–82 - PubMed
-
- Benner SA, Hutter D 2002. Phosphates, DNA, and the search for nonterrean life: A second generation model for genetic molecules. Bioorg Chem 30: 62–80 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources