

Review
doi: 10.1101/gad.1941710.
The Mdm2-p53 relationship evolves: Mdm2 swings both ways as an oncogene and a tumor suppressor
Affiliations
- PMID: 20679392
- PMCID: PMC2912554
- DOI: 10.1101/gad.1941710
Item in Clipboard
Review
The Mdm2-p53 relationship evolves: Mdm2 swings both ways as an oncogene and a tumor suppressor
Genes Dev.
.
Erratum in
- Genes Dev. 2010 Sep 15;24(18):2105
Abstract
Mdm2 has been well characterized as a negative regulator of the tumor suppressor p53. Recent studies have shown that Mdm2 is activated in response to a variety of oncogenic pathways independent of p53. Although its role as an oncogene via suppression of p53 function remains clear, growing evidence argues for p53-independent effects, as well as the remarkable possibility that Mdm2 has tumor suppressor functions in the appropriate context. Hence, Mdm2 is proving to be a key player in human cancer in its own right, and thus an important target for therapeutic intervention.
Figures
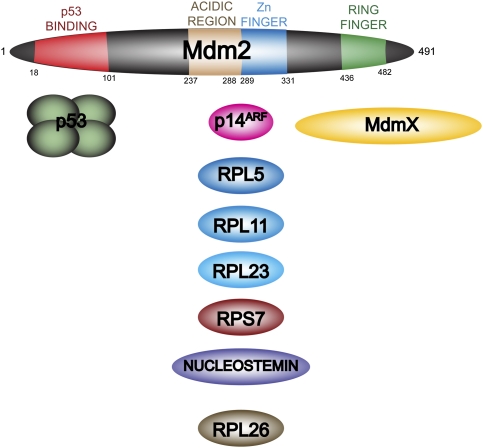
Mdm2 contains multiple protein-binding regions. Human Mdm2 consists of 491 amino acids. There are four previously characterized regions of the protein. At the N terminus, residues 18–101 are the main p53-binding region, and are the site that is targeted by several known Mdm2 inhibitors, including nutlin-3. In the central region is an acidic domain adjacent to a zinc finger (residues 237–331). This part of the protein interacts with a variety of regulatory factors, including the tumor suppressor p14ARF as well as multiple ribosomal or nucleolar proteins. The C terminus contains a RING finger that has been shown to be responsible for the E3 ubiquitin ligase activity, as well as the binding of the closely related MdmX.

Mdm2 regulates the p53-dependent response to DNA damage. Prior to DNA damage, Mdm2 interacts with both p53 and the ribosomal protein RPL26, leading to their ubiquitination and targeting for proteasomal degradation. Mdm2 bound to p53 has been localized to regulatory regions of a number of p53 target genes, leading to repression of their expression. Upon DNA damage, both Mdm2 and p53 become post-translationally modified such that they no longer interact. p53 is now capable of influencing gene expression. Recently, an additional mechanism for regulation of p53 expression by Mdm2 has been elucidated. It has been suggested that Mdm2 interacts with the mRNA encoding p53 itself and suppresses its translation. In addition, the ribosomal protein RPL26 has been shown to be an obligate effector of p53 translation as well. Thus, upon DNA damage, the interaction of RPL26 with Mdm2 is also disrupted, leading to increased levels and its association with p53 mRNA, facilitating its translation and increasing p53 levels.

The interaction of Mdm2 and p53 is disrupted in response to oncogenic activation as well as ribosomal stress. The interaction of Mdm2 with p53 is also disrupted via the binding of specific regulatory proteins. In response to oncogenic activation, p14ARF interacts with the central region of Mdm2, thereby inhibiting its activity toward p53. Nucleolar or ribosomal stress has been suggested to lead to the release of several proteins that also interact with Mdm2, preventing it from negatively regulating p53.

Mdm2 is transcriptionally regulated by multiple pathways. (A) There are two promoters that function in the regulation of Mdm2 expression. The P1 promoter controls basal expression of Mdm2 and is upstream of exon 1a. The P2 promoter is further downstream and uses a start site at an alternate first exon, 1b. The P2 promoter is inducible, and is regulated by response elements for a variety of transcription factors, including RXR, AP-1, the Ets family, Smad2, and Smad3, as well as p53 itself. A cluster of nGGGGC boxes act downstream from the Erk pathway. Three GC boxes control expression of Mdm2, with one of these containing the site of a single-nucleotide polymorphism (SNP309) that influences the binding of the Sp-1 transcription factor. (B) The two transcripts from the P1 and P2 promoters have distinct 5′ untranslated regions (encoded by either exon 1a or exon 1b) and are different lengths. It should be noted that sequences corresponding to exon 1b are not present in the mature P1 transcript. Nevertheless, the start site of translation is contained in a shared exon 2. Thus, the Mdm2 protein that is expressed is identical between the two transcripts. There has been a suggestion, however, that further splicing of the transcript may be influenced by promoter selection.

Oncogenic pathways enhance Mdm2 activity. Growth factor signaling via tyrosine kinase receptors and Ras lead to the activation of AP-1 and Ets family members, which have been shown to transcriptionally up-regulate Mdm2. TGFβ signaling has also been shown to increase Mdm2 via the Smad transcription factors. Phosphatidyl-3 kinase (PI3K) signaling through the Akt kinase leads to phosphorylation of Mdm2 on sites distinct from those modified in response to DNA damage. This results in nuclear import of Mdm2 and enhancement of its ability to inhibit p53 (Zhou et al. 2001).

Tumor suppressor pathways control Mdm2 function. The p14ARF protein is a negative regulator of Mdm2 function and is transcriptionally controlled by the E2F family. The RB pathway regulates E2F function. Multiple components of this pathway have been shown to play a role in human cancer as either oncogenes (Cyclin D, Cdk4, and Cdk6) or tumor suppressors (p16INK4A and pRB). TGFβ signaling can also exert a suppressor effect via transcriptional up-regulation of the cyclin-dependent kinase inhibitor p15INK4B. It should also be noted that TGFβ signaling has been implicated in regulating p14ARF expression more directly via the direct interaction of Smad proteins with the ARF genomic locus (Freeman-Anderson et al. 2009).
References
-
- Barak Y, Gottlieb E, Juven-Gershon T, Oren M 1994. Regulation of mdm2 expression by p53: Alternative promoters produce transcripts with nonidentical translation potential. Genes Dev 8: 1739–1749 - PubMed
-
- Bartel F, Taubert H, Harris LC 2002. Alternative and aberrant splicing of MDM2 mRNA in human cancer. Cancer Cell 2: 9–15 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous