

Sodium-calcium exchange is essential for effective triggering of calcium release in mouse heart
- PMID: 20682252
- PMCID: PMC2913203
- DOI: 10.1016/j.bpj.2010.04.071
Sodium-calcium exchange is essential for effective triggering of calcium release in mouse heart
Abstract
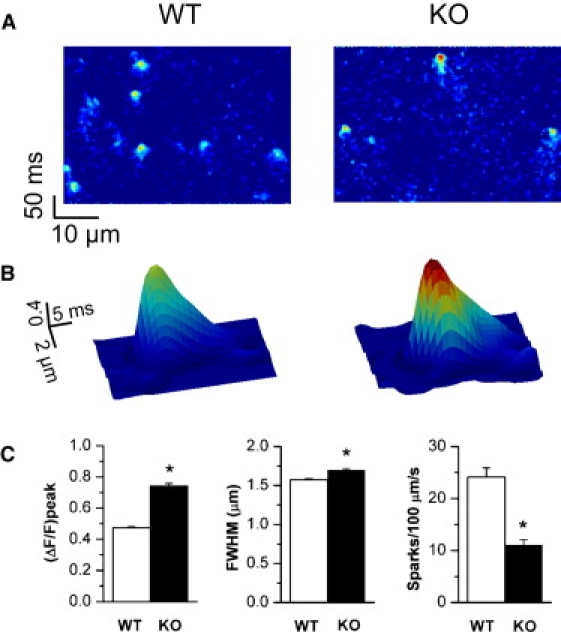
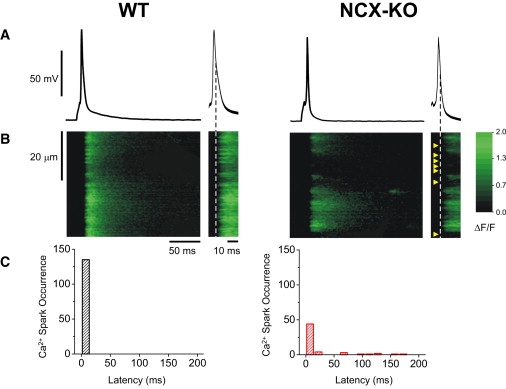
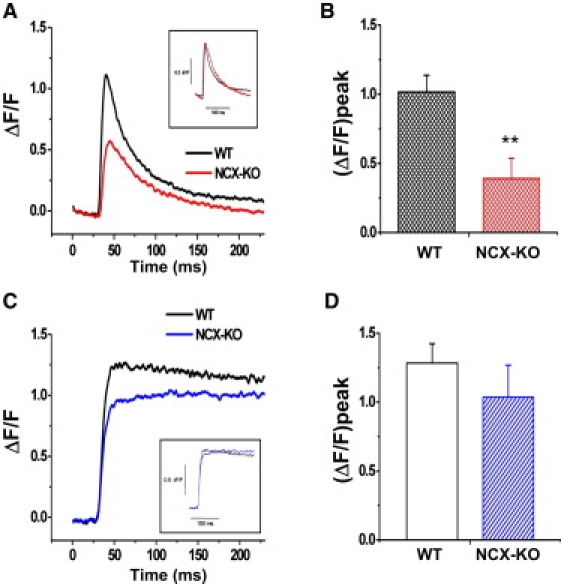
In cardiac myocytes, excitation-contraction coupling depends upon sarcoplasmic reticular Ca2+ release triggered by Ca2+ influx through L-type Ca2+ channels. Although Na+-Ca2+ exchange (NCX) is essential for Ca2+ extrusion, its participation in the trigger process of excitation-contraction coupling is controversial. To investigate the role of NCX in triggering, we examined Ca2+ sparks in ventricular cardiomyocytes isolated from wild-type (WT) and cardiac-specific NCX knockout (KO) mice. Myocytes from young NCX KO mice are known to exhibit normal resting cytosolic Ca2+ and normal Ca2+ transients despite reduced L-type Ca2+ current. We loaded myocytes with fluo-3 to image Ca2+ sparks using confocal microscopy in line-scan mode. The frequency of spontaneous Ca2+ sparks was reduced in KO myocytes compared with WT. However, spark amplitude and width were increased in KO mice. Permeabilizing the myocytes with saponin eliminated differences between spontaneous sparks in WT and KO mice. These results suggest that sarcolemmal processes are responsible for the reduced spark frequency and increased spark width and amplitude in KO mice. When myocytes were loaded with 1 mM fluo-3 and 3 mM EGTA via the patch pipette to buffer diadic cleft Ca2+, the number of sparks triggered by action potentials was reduced by 60% in KO cells compared to WT cells, despite similar SR Ca2+ content in both cell types. When EGTA was omitted from the pipette solution, the number of sparks triggered in KO and WT myocytes was similar. Although the number of sparks was restored in KO cells, Ca2+ release was asynchronous. These results suggest that high subsarcolemmal Ca2+ is required to ensure synchronous triggering with short spark latency in the absence of NCX. In WT mice, high subsarcolemmal Ca2+ is not required for synchronous triggering, because NCX is capable of priming the diadic cleft with sufficient Ca2+ for normal triggering, even when subsarcolemmal Ca(2+) is lowered by EGTA. Thus, reducing subsarcolemmal Ca2+ with EGTA in NCX KO mice reveals the dependence of Ca2+ release on NCX.
2010 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures



Similar articles
-
Heterogeneity of transverse-axial tubule system in mouse atria: Remodeling in atrial-specific Na+-Ca2+ exchanger knockout mice.J Mol Cell Cardiol. 2017 Jul;108:50-60. doi: 10.1016/j.yjmcc.2017.05.008. Epub 2017 May 19. J Mol Cell Cardiol. 2017. PMID: 28529049 Free PMC article.
-
Ca2+ sparks induced by Na/Ca exchange.Cell Calcium. 2003 Jul;34(1):11-7. doi: 10.1016/s0143-4160(03)00017-4. Cell Calcium. 2003. PMID: 12767888
-
Excitation-contraction coupling in Na+-Ca2+ exchanger knockout mice: reduced transsarcolemmal Ca2+ flux.Circ Res. 2005 Dec 9;97(12):1288-95. doi: 10.1161/01.RES.0000196563.84231.21. Epub 2005 Nov 17. Circ Res. 2005. PMID: 16293789 Free PMC article.
-
Cardiac sodium-calcium exchange and efficient excitation-contraction coupling: implications for heart disease.Adv Exp Med Biol. 2013;961:355-64. doi: 10.1007/978-1-4614-4756-6_30. Adv Exp Med Biol. 2013. PMID: 23224894 Free PMC article. Review.
-
Na/Ca exchange and contraction of the heart.J Mol Cell Cardiol. 2013 Aug;61:28-33. doi: 10.1016/j.yjmcc.2013.06.001. Epub 2013 Jun 12. J Mol Cell Cardiol. 2013. PMID: 23770352 Free PMC article. Review.
Cited by
-
The Role of Saponins in the Treatment of Neuropathic Pain.Molecules. 2022 Jun 20;27(12):3956. doi: 10.3390/molecules27123956. Molecules. 2022. PMID: 35745079 Free PMC article. Review.
-
Altered Intracellular Calcium Homeostasis and Arrhythmogenesis in the Aged Heart.Int J Mol Sci. 2019 May 14;20(10):2386. doi: 10.3390/ijms20102386. Int J Mol Sci. 2019. PMID: 31091723 Free PMC article. Review.
-
Subcellular heterogeneity of ryanodine receptor properties in ventricular myocytes with low T-tubule density.PLoS One. 2011;6(10):e25100. doi: 10.1371/journal.pone.0025100. Epub 2011 Oct 13. PLoS One. 2011. PMID: 22022376 Free PMC article.
-
Downregulation of FKBP5 Promotes Atrial Arrhythmogenesis.Circ Res. 2023 Jun 23;133(1):e1-e16. doi: 10.1161/CIRCRESAHA.122.322213. Epub 2023 May 8. Circ Res. 2023. PMID: 37154033 Free PMC article.
-
Na(+)/K)+)-ATPase α2-isoform preferentially modulates Ca2(+) transients and sarcoplasmic reticulum Ca2(+) release in cardiac myocytes.Cardiovasc Res. 2012 Sep 1;95(4):480-6. doi: 10.1093/cvr/cvs213. Epub 2012 Jun 27. Cardiovasc Res. 2012. PMID: 22739122 Free PMC article.
References
-
- Fabiato A. Calcium-induced release of calcium from the cardiac sarcoplasmic reticulum. Am. J. Physiol. 1983;245:C1–C14. - PubMed
-
- Izu L.T., Balke C.W. The Ca2+ synapse redo: a matter of location, location, location. Circ. Res. 2002;91:276–277. - PubMed
-
- Cheng H., Lederer W.J., Canneldl M.B. Calcium sparks: elementary events underlying excitation-contraction coupling in heart muscle. Science. 1993;262:740–744. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
