

Robust IgA and IgG-producing antibody forming cells in the diffuse-NALT and lungs of Sendai virus-vaccinated cotton rats associate with rapid protection against human parainfluenza virus-type 1
- PMID: 20682364
- PMCID: PMC2950074
- DOI: 10.1016/j.vaccine.2010.07.068
Robust IgA and IgG-producing antibody forming cells in the diffuse-NALT and lungs of Sendai virus-vaccinated cotton rats associate with rapid protection against human parainfluenza virus-type 1
Abstract
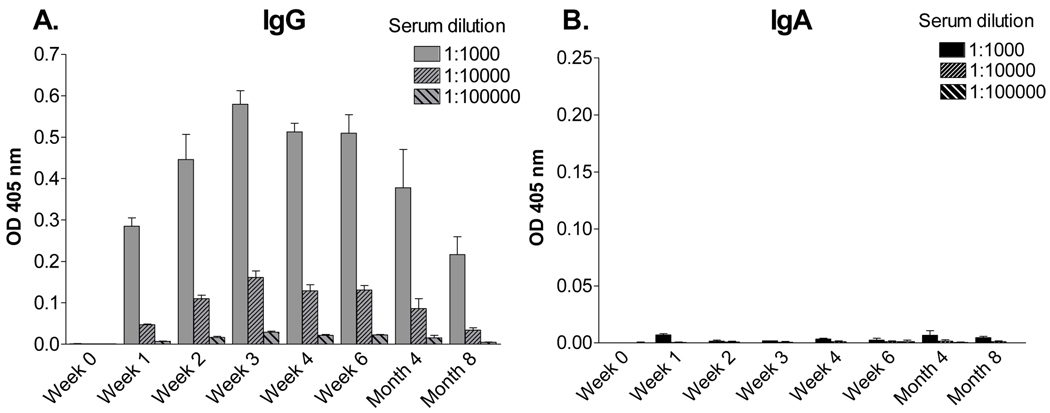
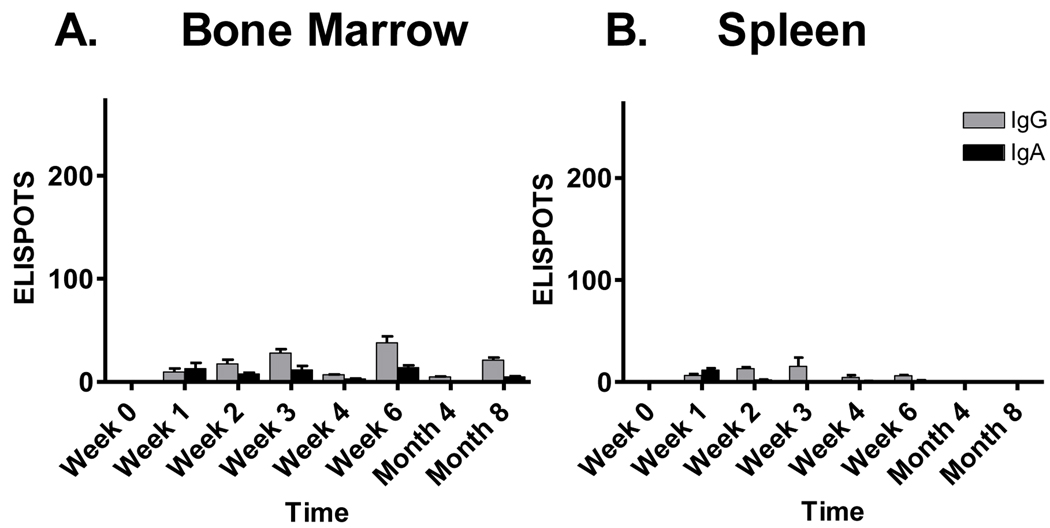
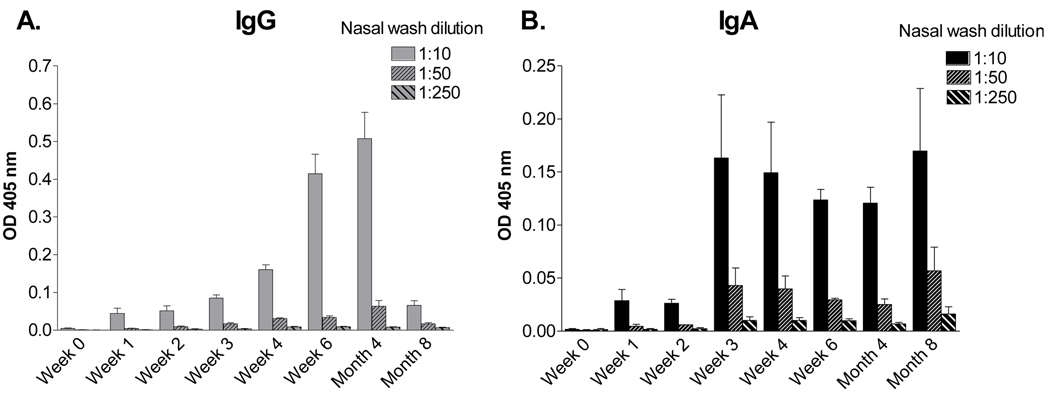
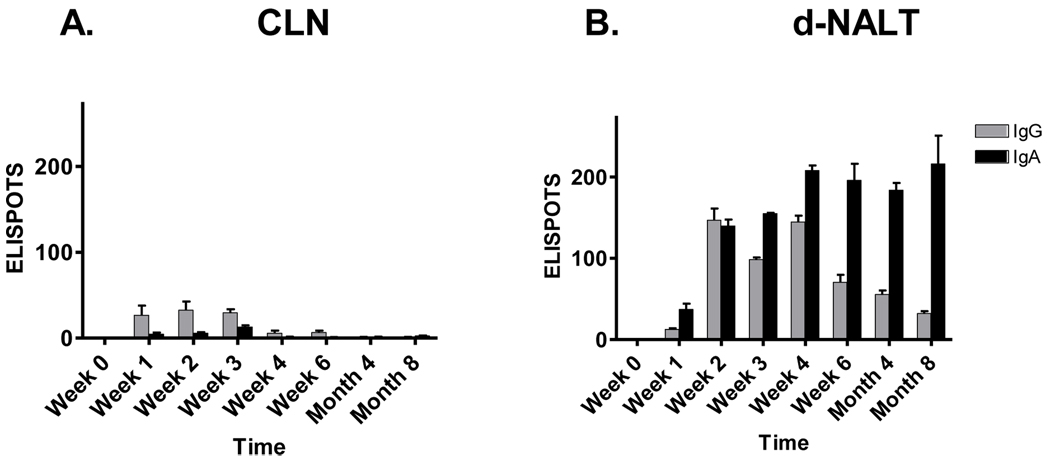
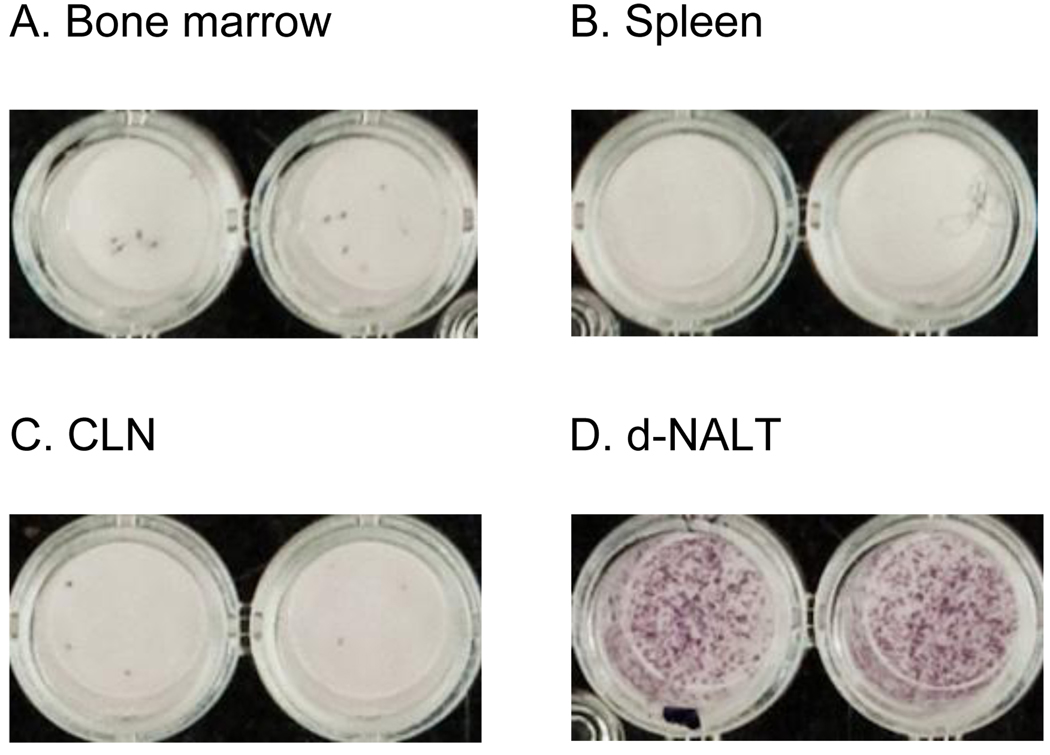
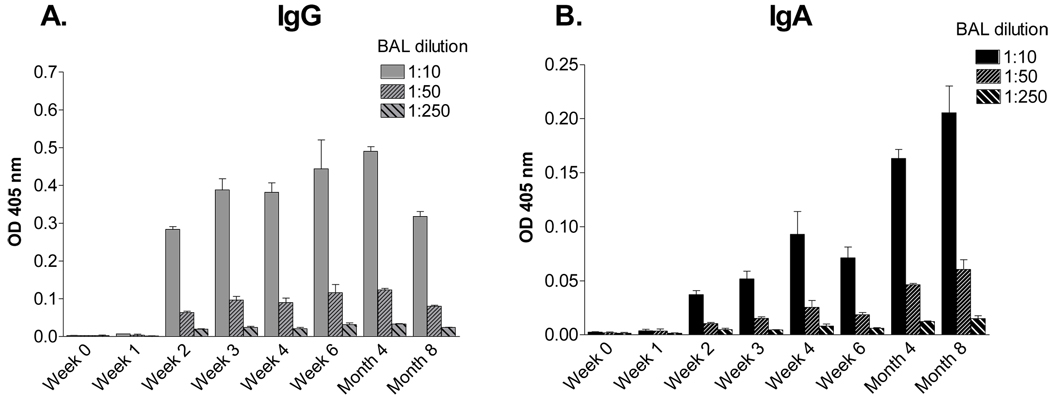
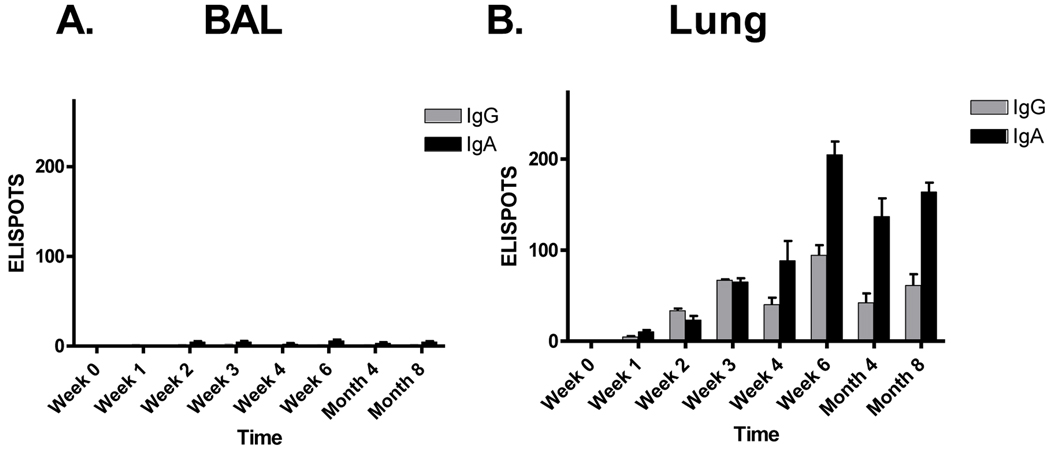
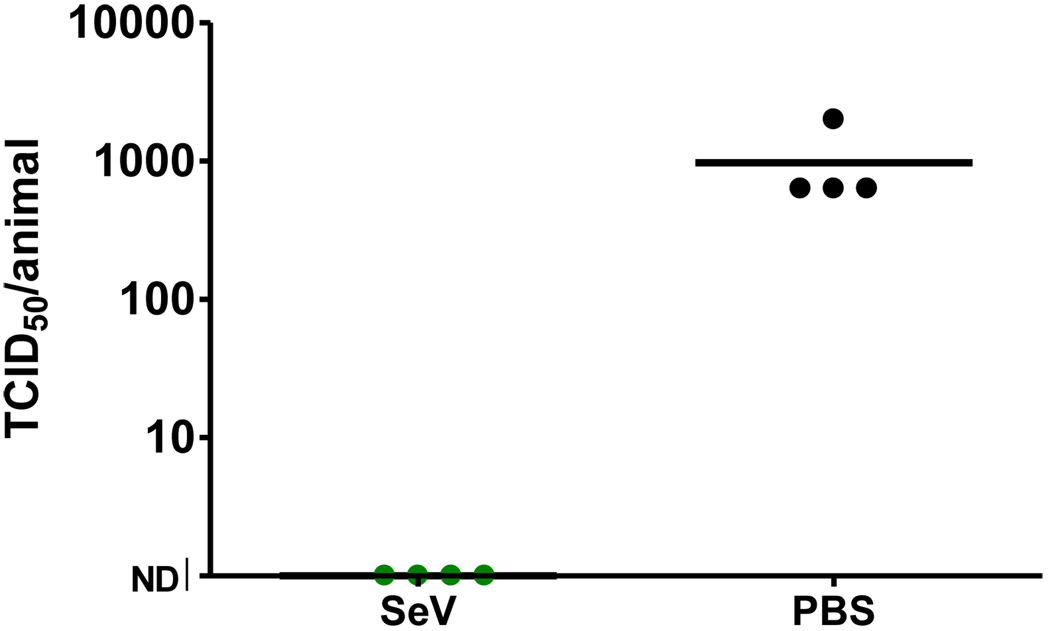
Sendai virus (SeV), a natural mouse pathogen, shows considerable promise as a candidate vaccine for human parainfluenza virus-type 1 (hPIV-1), and also as a vaccine vector for other serious pathogens of infants including respiratory syncytial virus (RSV). In an effort to define correlates of immunity, we examined the virus-specific serum antibody of cotton rats inoculated intranasally (I.N.) with SeV. Virus-specific antibody forming cells (AFCs) were also measured in the bone marrow, because these are considered responsible for durable serum antibody levels in other viral systems. Results showed that a single SeV inoculation was sufficient to induce virus-specific serum antibodies and bone marrow-resident AFCs that persisted for as many as 8 months post-vaccination. Given that the predominant SeV-specific serum antibody isotype was IgG, an isotype that traffics poorly to the upper respiratory tract (URT), we asked if local nasal and lung-associated antibodies and AFCs were also present. Studies showed that: (i) SeV-specific antibodies appeared in the URT and lower respiratory tract (LRT) within 7 days after immunization, (ii) corresponding AFCs were present in the diffuse-NALT (d-NALT) and lung, (iii) AFCs in the d-NALT and lung peaked at approximately 6 weeks and persisted for the lifetime of the animal, reaching a level exceeding that of the bone marrow by an order of magnitude, (iv) IgA was the dominant isotype among AFCs in the d-NALT and lung at 4-weeks post-vaccination and thereafter, and (v) antibody and AFC responses associated with the prevention of lung infection when animals were challenged with hPIV-1 just 1 week after vaccination.
Copyright © 2010 Elsevier Ltd. All rights reserved.
Figures
Similar articles
-
Vitamin A deficiency disrupts vaccine-induced antibody-forming cells and the balance of IgA/IgG isotypes in the upper and lower respiratory tract.Viral Immunol. 2012 Aug;25(4):341-4. doi: 10.1089/vim.2012.0023. Epub 2012 Jul 19. Viral Immunol. 2012. PMID: 22813425 Free PMC article.
-
Sendai virus recombinant vaccine expressing hPIV-3 HN or F elicits protective immunity and combines with a second recombinant to prevent hPIV-1, hPIV-3 and RSV infections.Vaccine. 2008 Jun 25;26(27-28):3480-8. doi: 10.1016/j.vaccine.2008.04.022. Epub 2008 May 1. Vaccine. 2008. PMID: 18499307 Free PMC article.
-
Phenotypes and functions of persistent Sendai virus-induced antibody forming cells and CD8+ T cells in diffuse nasal-associated lymphoid tissue typify lymphocyte responses of the gut.Virology. 2011 Feb 20;410(2):429-436. doi: 10.1016/j.virol.2010.12.017. Epub 2011 Jan 11. Virology. 2011. PMID: 21227475 Free PMC article.
-
Sendai virus as a backbone for vaccines against RSV and other human paramyxoviruses.Expert Rev Vaccines. 2016;15(2):189-200. doi: 10.1586/14760584.2016.1114418. Epub 2015 Dec 9. Expert Rev Vaccines. 2016. PMID: 26648515 Free PMC article. Review.
-
Sendai Virus-Vectored Vaccines That Express Envelope Glycoproteins of Respiratory Viruses.Viruses. 2021 May 29;13(6):1023. doi: 10.3390/v13061023. Viruses. 2021. PMID: 34072332 Free PMC article. Review.
Cited by
-
Intranasal Sendai Virus Vaccination of Seropositive Children 1 to 2 Years of Age in a Phase I Clinical Trial Boosts Immune Responses Toward Human Parainfluenza Virus Type 1.Vaccines (Basel). 2025 Apr 19;13(4):430. doi: 10.3390/vaccines13040430. Vaccines (Basel). 2025. PMID: 40333329 Free PMC article.
-
Clonally related CD8+ T cells responsible for rapid population of both diffuse nasal-associated lymphoid tissue and lung after respiratory virus infection.J Immunol. 2011 Jul 15;187(2):835-41. doi: 10.4049/jimmunol.1100125. Epub 2011 Jun 20. J Immunol. 2011. PMID: 21690324 Free PMC article.
-
Intranasal immunization with an RBD-hemagglutinin fusion protein harnesses preexisting immunity to enhance antigen-specific responses.J Clin Invest. 2023 Dec 1;133(23):e166827. doi: 10.1172/JCI166827. J Clin Invest. 2023. PMID: 38038133 Free PMC article.
-
Relationships among dissemination of primary parainfluenza virus infection in the respiratory tract, mucosal and peripheral immune responses, and protection from reinfection: a noninvasive bioluminescence-imaging study.J Virol. 2015 Apr;89(7):3568-83. doi: 10.1128/JVI.03581-14. Epub 2015 Jan 14. J Virol. 2015. PMID: 25589649 Free PMC article.
-
CD4+ T cells support establishment of RSV-specific IgG and IgA antibody secreting cells in the upper and lower murine respiratory tract following RSV infection.Vaccine. 2017 May 9;35(20):2617-2621. doi: 10.1016/j.vaccine.2017.03.073. Epub 2017 Apr 11. Vaccine. 2017. PMID: 28410812 Free PMC article.
References
-
- Henderson FW. Pulmonary infections with respiratory syncytial virus and the parainfluenza viruses. Semin Respir Infect. 1987 Jun;2(2):112–121. - PubMed
-
- Henrickson KJ, Kuhn SM, Savatski LL. Epidemiology and cost of infection with human parainfluenza virus types 1 and 2 in young children. Clin Inf Dis. 1994;18:770–779. - PubMed
-
- Henrickson KJ, Hoover S, Kehl KS, Hua W. National disease burden of respiratory viruses detected in children by polymerase chain reaction. Pediatr Infect DIs J. 2004 Jan;23(1 Suppl):S11–S18. - PubMed
-
- Heilman CA. From the National Institute of Allergy and Infectious Diseases and the World Health Organization. Respiratory syncytial and parainfluenza viruses. J Infect Dis. 1990 Mar;161(3):402–406. - PubMed
-
- Ishida N, Homma M. Sendai virus. Adv Virus Res. 1978;23:349–383. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
