

Sorafenib activates CD95 and promotes autophagy and cell death via Src family kinases in gastrointestinal tumor cells
- PMID: 20682655
- PMCID: PMC2933415
- DOI: 10.1158/1535-7163.MCT-10-0274
Sorafenib activates CD95 and promotes autophagy and cell death via Src family kinases in gastrointestinal tumor cells
Abstract
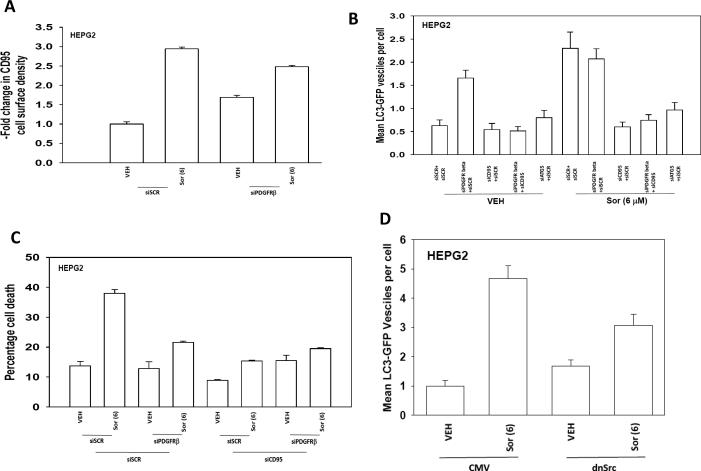
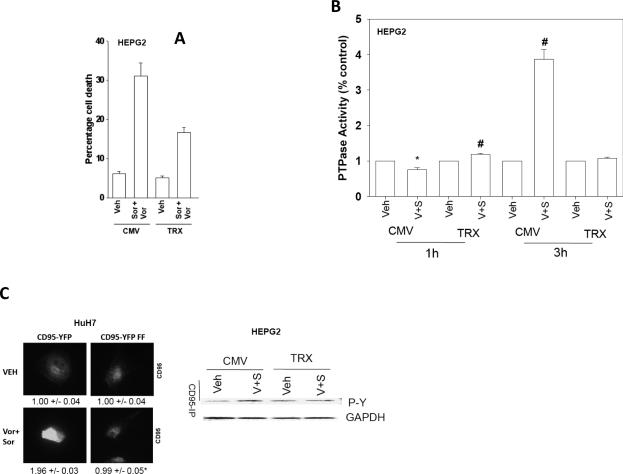
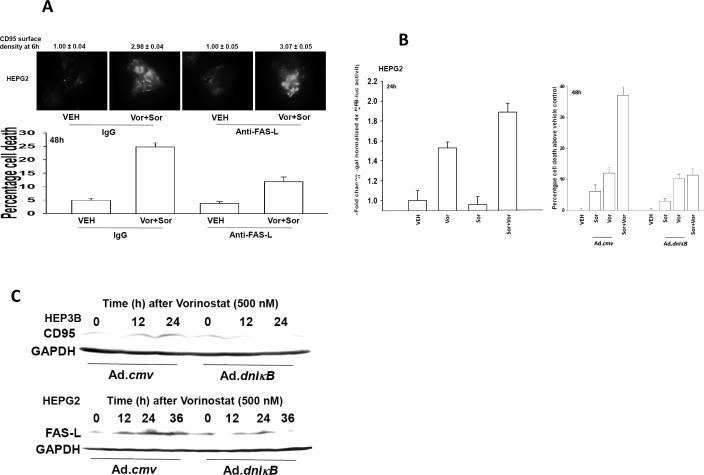
Sorafenib and vorinostat interact in a synergistic fashion to kill carcinoma cells by activating CD95; the present studies have determined how sorafenib and vorinostat individually contribute to CD95 activation. Sorafenib (3-6 micromol/L) promoted a dose-dependent increase in Src Y416, ERBB1 Y845 and CD95 Y232/Y291 phosphorylation, and Src Y527 dephosphorylation. Low levels of sorafenib-induced (3 micromol/L) CD95 tyrosine phosphorylation did not promote surface localization whereas sorafenib (6 micromol/L), or sorafenib (3 micromol/L) and vorinostat (500 nmol/L) treatment promoted higher levels of CD95 phosphorylation which correlated with DISC formation, receptor surface localization, and autophagy. CD95 (Y232F, Y291F) was not tyrosine phosphorylated and was unable to localize plasma membrane or induce autophagy. Knockdown/knockout of Src family kinases abolished sorafenib-induced CD95 tyrosine phosphorylation, DISC formation, and the induction of cell death and autophagy. Knockdown of platelet-ived growth factor receptor-beta enhanced Src Y416 and CD95 tyrosine phosphorylation, which correlated with elevated CD95 plasma membrane levels and autophagy, and with a reduced ability of sorafenib to promote CD95 membrane localization. Vorinostat increased reactive oxygen species levels, and in a delayed NF kappa B-dependent fashion, those of FAS ligand and CD95. Neutralization of FAS-L did not alter the initial rapid drug-induced activation of CD95; however, neutralization of FAS-L reduced sorafenib + vorinostat toxicity by approximately 50%. Thus, sorafenib contributes to CD95 activation by promoting receptor tyrosine phosphorylation, whereas vorinostat contributes to CD95 activation via the initial facilitation of reactive oxygen species generation and subsequently of FAS-L expression.
(c) 2010 AACR.
Figures



References
-
- Parkin DM, Bray F, Ferlay J, Pisani P. Global cancer statistics. CA Cancer J Clin 2005. 2002;55:74–108. - PubMed
-
- Bilimoria KY, Bentrem DJ, Lillemoe KD, Talamonti MS, Ko CY. Pancreatic Cancer Quality Indicator Development Expert Panel, American College of Surgeons. Assessment of pancreatic cancer care in the United States based on formally developed quality indicators. J Natl Cancer Inst. 2009;101:848–59. - PMC - PubMed
-
- Dent P. MAP kinase pathways in the control of hepatocyte growth, metabolism and survival. In: Dufour JF, Clavien P-A, editors. Signaling Pathways in Liver Diseases. Springer Press; 2005. pp. 223–238. Chapter 19.
-
- Dent P, Yacoub A, Fisher PB, Hagan MP, Grant S. MAPK pathways in radiation responses. Oncogene. 2003;22:5885–96. - PubMed
-
- Valerie K, Yacoub A, Hagan MP, et al. Radiation-induced cell signaling: inside-out and outside-in. Mol Cancer Ther. 2007;6:789–801. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 CA108520/CA/NCI NIH HHS/United States
- R01-CA77141/CA/NCI NIH HHS/United States
- R01-CA63753/CA/NCI NIH HHS/United States
- R01 CA063753/CA/NCI NIH HHS/United States
- R01-DK52825/DK/NIDDK NIH HHS/United States
- R01 CA093738/CA/NCI NIH HHS/United States
- R01 CA141703/CA/NCI NIH HHS/United States
- P01 CA104177/CA/NCI NIH HHS/United States
- R01 DK052825/DK/NIDDK NIH HHS/United States
- P01-CA104177/CA/NCI NIH HHS/United States
- R01-CA108520/CA/NCI NIH HHS/United States
- R01-CA93738/CA/NCI NIH HHS/United States
- R01 CA150214/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
