

The p38 SAPK is recruited to chromatin via its interaction with transcription factors
- PMID: 20682780
- PMCID: PMC2951253
- DOI: 10.1074/jbc.M110.155846
The p38 SAPK is recruited to chromatin via its interaction with transcription factors
Abstract
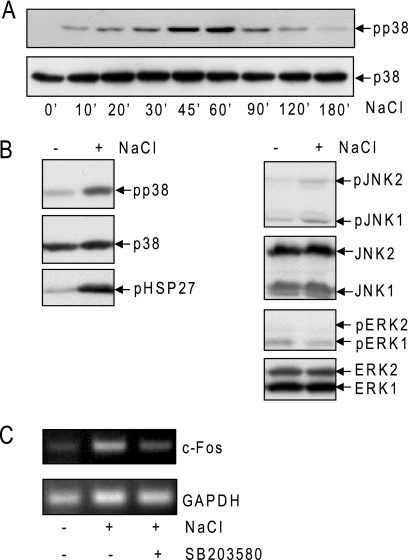
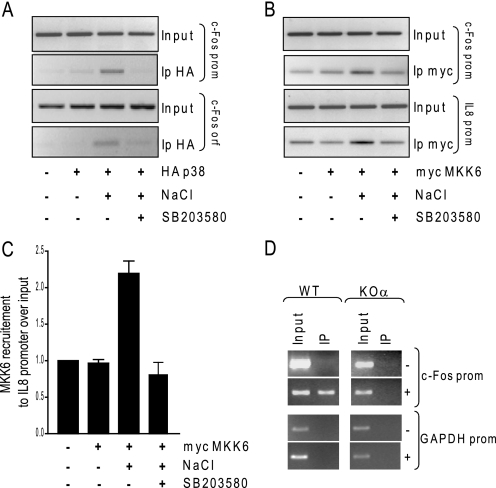
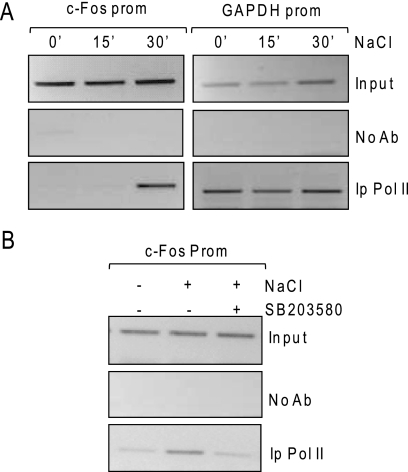
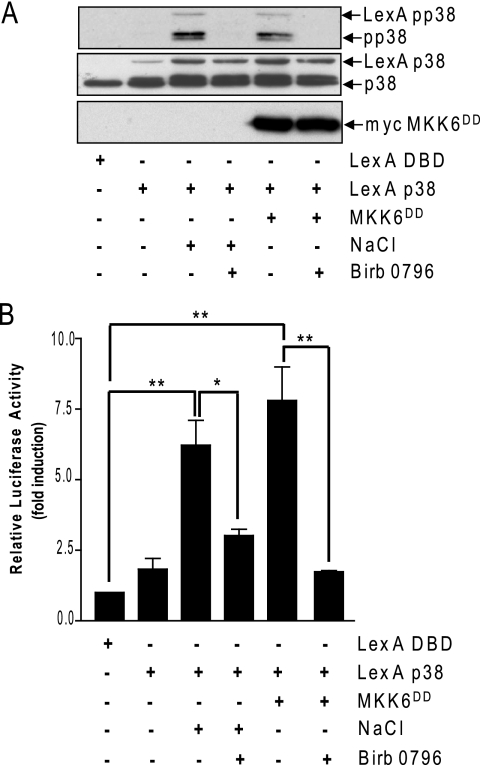
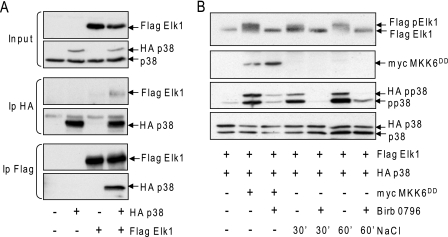
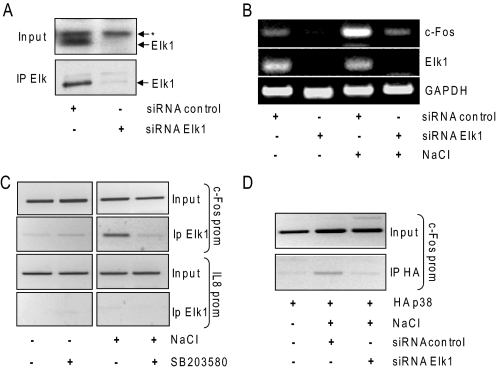
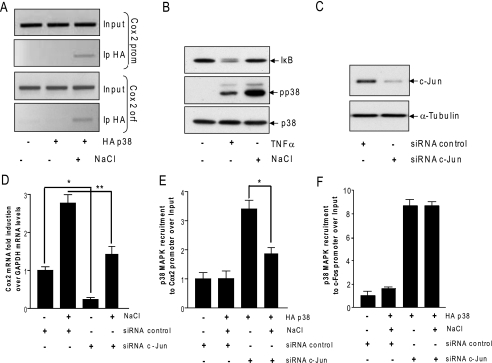
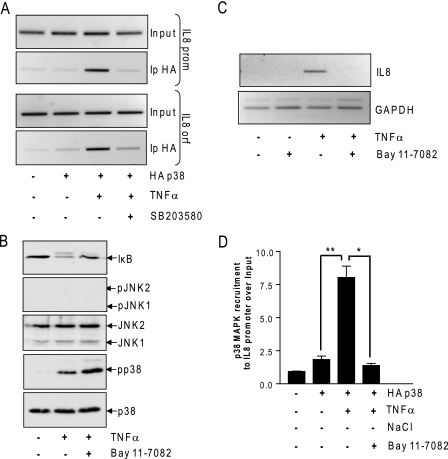
In mammals, the stress-activated protein kinase (SAPK) p38 coordinates a rapid and complex transcriptional program to adapt to sudden changes in the extracellular environment. Although a number of genes have been reported to be under the control of p38, the basic mechanisms of transcriptional regulation by this SAPK remain uncharacterized. Here we show that in response to osmotic shock, anisomycin- or TNFα-activated p38 SAPK is recruited to stress-induced genes. The MAPKK MKK6 is also found at stress-responsive promoters. The recruitment of RNA polymerase II complex to the target promoters requires p38 activity. Moreover, when tethered to DNA as a LexA fusion protein, p38 activates transcription in a stress-regulated manner. Thus, p38 activity allows for recruitment of RNA polymerase and transcription initiation. p38 directly phosphorylates and interacts with the transcription factor Elk1. p38 activity is necessary for the recruitment of Elk1 to the c-Fos promoter, and knocking down Elk1 by siRNAs compromises both p38 recruitment to the c-Fos promoter and c-Fos transcriptional up-regulation upon osmostress. In addition, p38 recruitment to the osmoinducible gene Cox2 and the TNFα target gene IL8 is mediated by the transcription factors AP1 and NFκB, respectively. Therefore, anchoring of active SAPK to target genes is mediated by transcription factors. The presence of active p38 at open reading frames also suggests the involvement of the SAPK in elongation. Taken together, SAPK recruitment to target genes appears to be a broad mechanism to regulate transcription that has been preserved from yeast to mammals.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
