

Interaction of ibogaine with human alpha3beta4-nicotinic acetylcholine receptors in different conformational states
- PMID: 20684041
- PMCID: PMC4609575
- DOI: 10.1016/j.biocel.2010.05.011
Interaction of ibogaine with human alpha3beta4-nicotinic acetylcholine receptors in different conformational states
Abstract
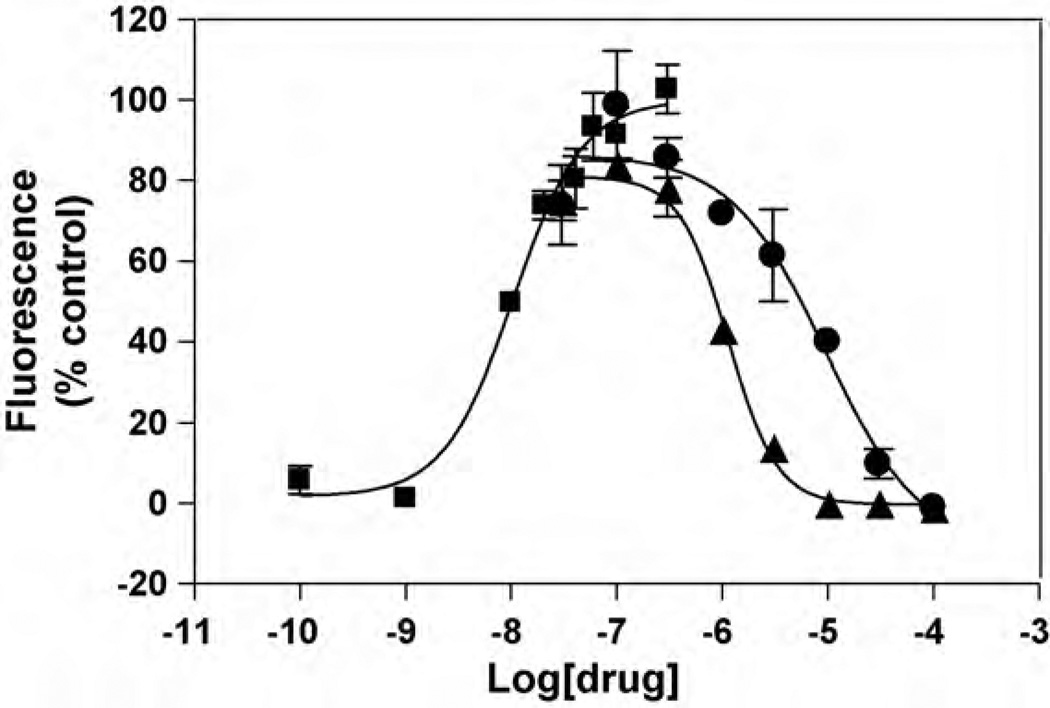
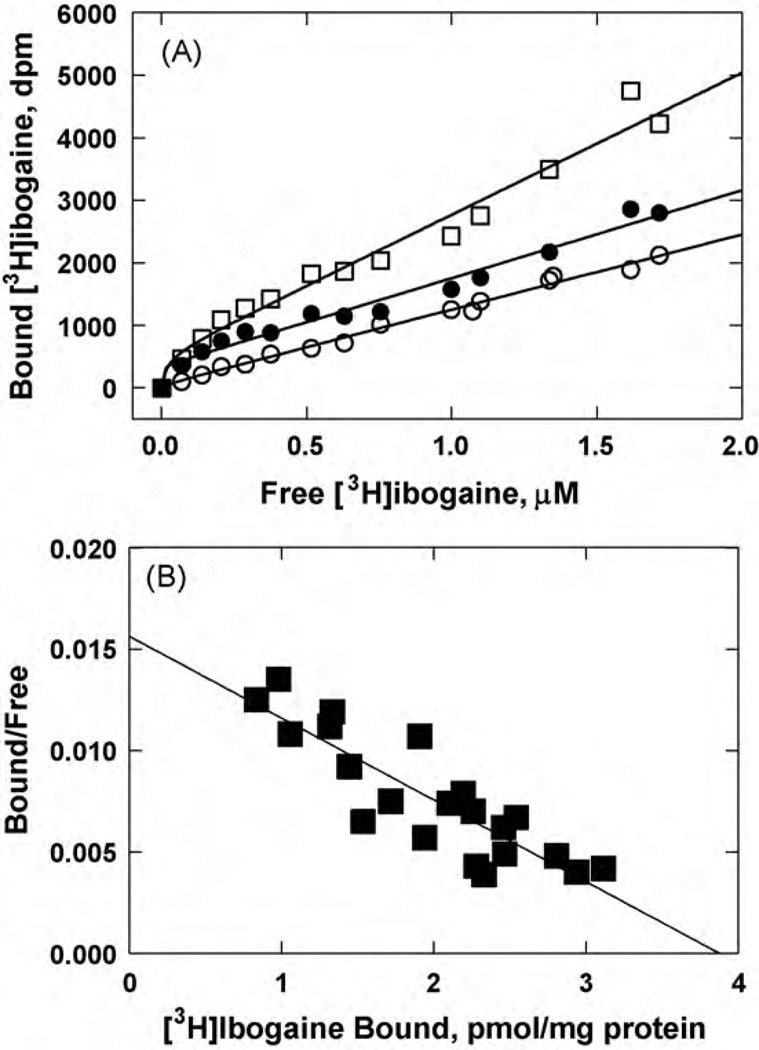
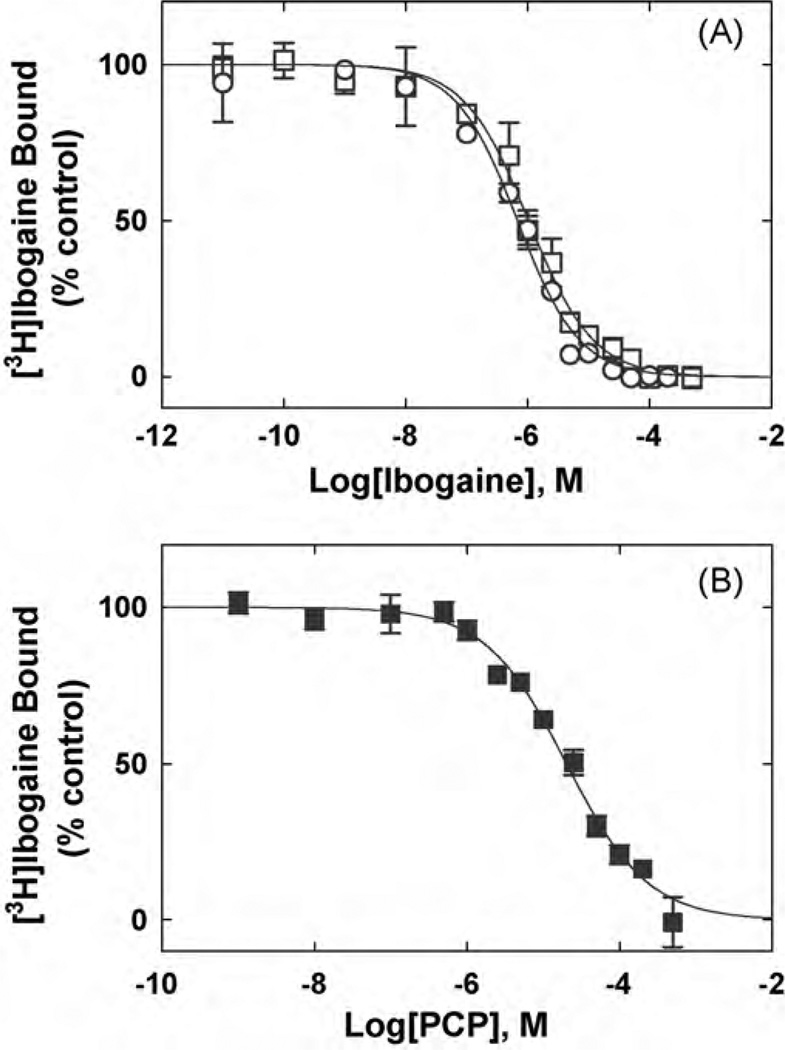
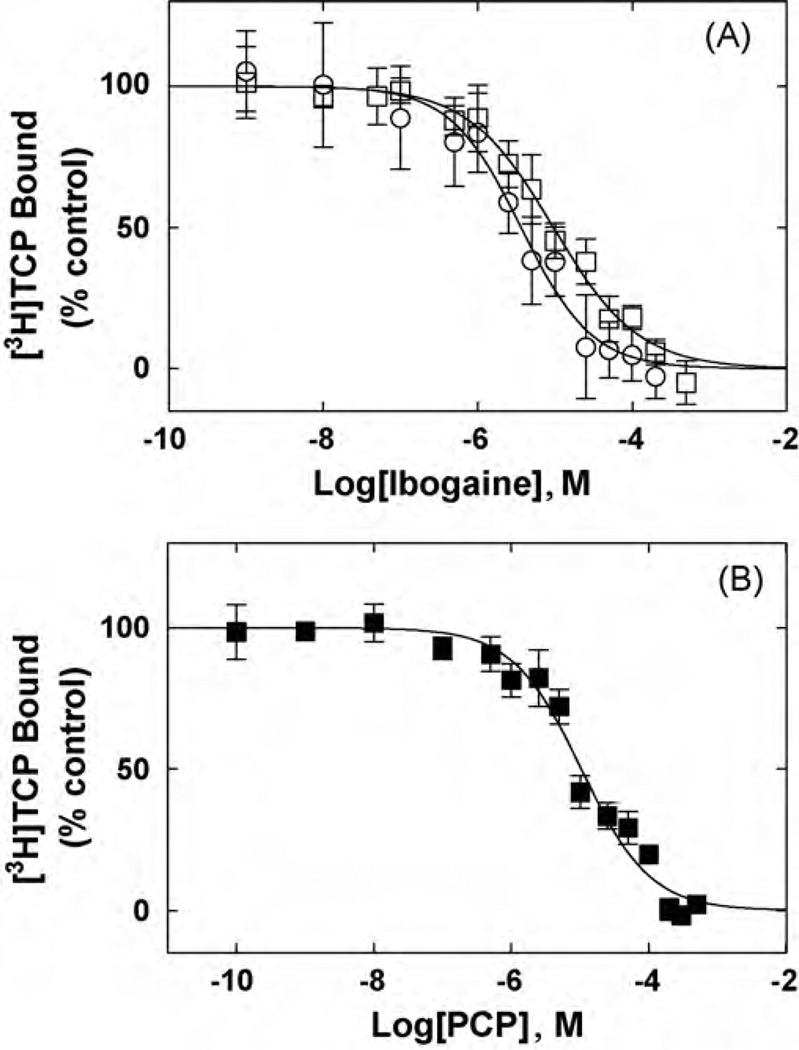
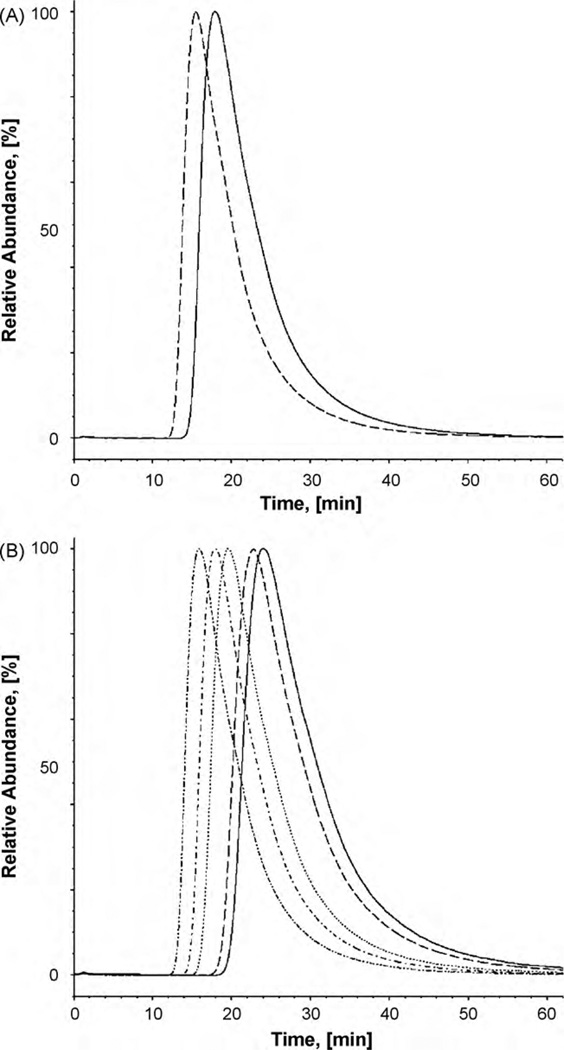
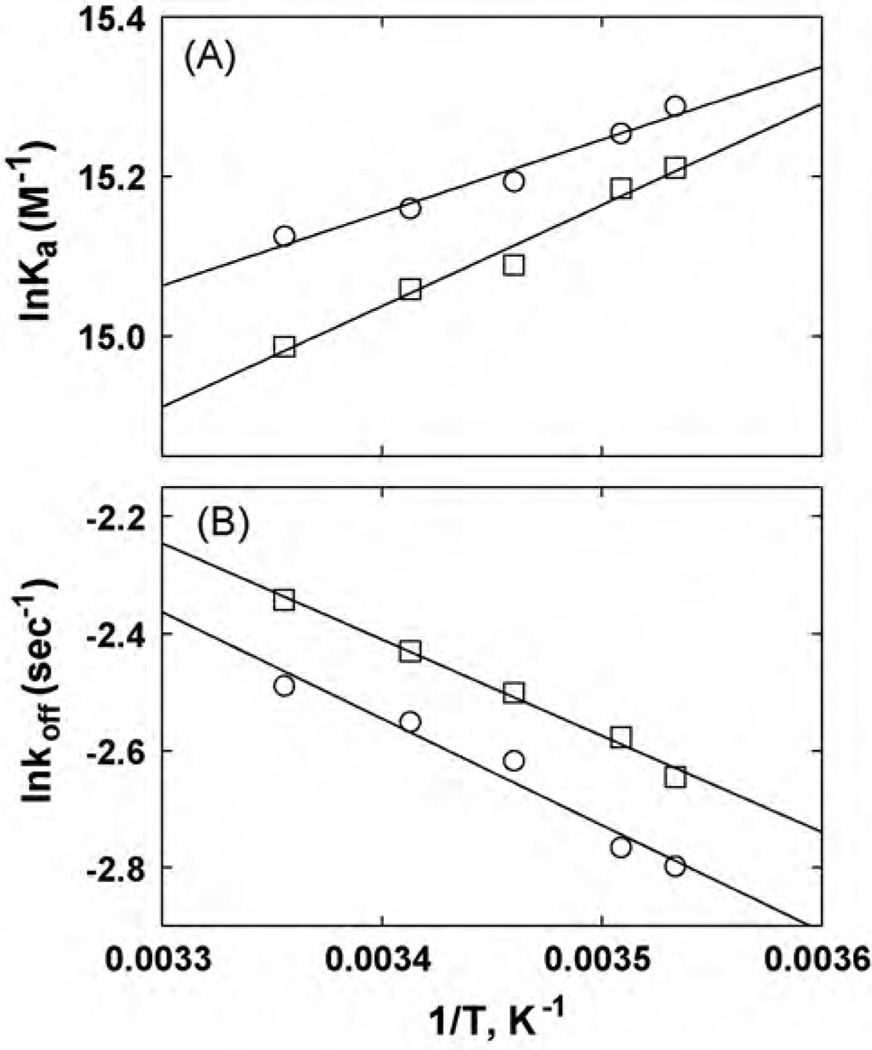
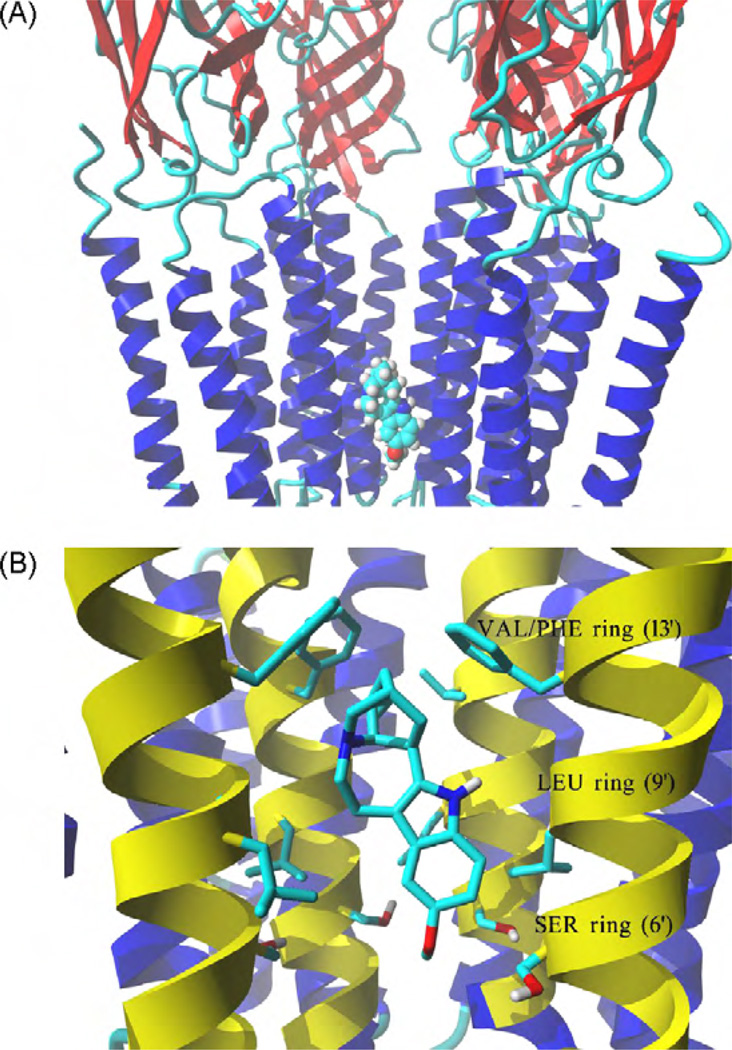
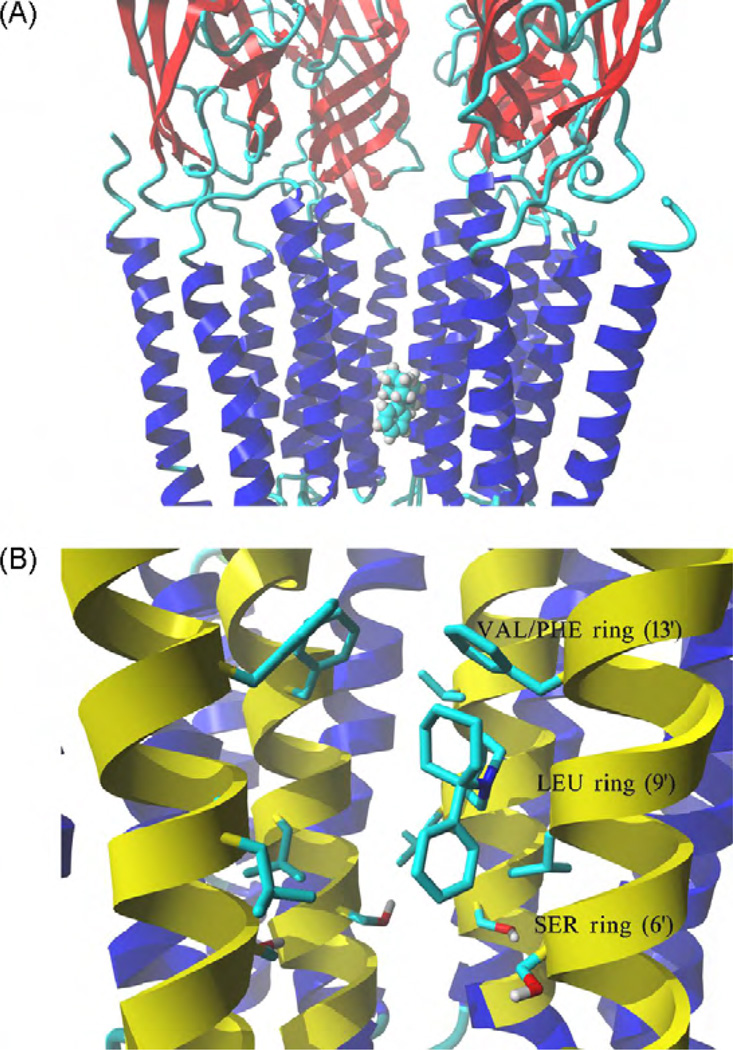
The interaction of ibogaine and phencyclidine (PCP) with human (h) alpha3beta4-nicotinic acetylcholine receptors (AChRs) in different conformational states was determined by functional and structural approaches including, radioligand binding assays, Ca2+ influx detections, and thermodynamic and kinetics measurements. The results established that (a) ibogaine inhibits (+/-)-epibatidine-induced Ca2+ influx in h(alpha)3beta4 AChRs with approximately 9-fold higher potency than that for PCP, (b) [3H]ibogaine binds to a single site in the h(alpha)3beta4 AChR ion channel with relatively high affinity (Kd = 0.46 +/- 0.06 microM), and ibogaine inhibits [3H]ibogaine binding to the desensitized h(alpha)3beta4 AChR with slightly higher affinity compared to the resting AChR. This is explained by a slower dissociation rate from the desensitized ion channel compared to the resting ion channel, and (c) PCP inhibits [3H]ibogaine binding to the h(alpha)3beta4 AChR, suggesting overlapping sites. The experimental results correlate with the docking simulations suggesting that ibogaine and PCP interact with a binding domain located between the serine (position 6') and valine/phenylalanine (position 13') rings. This interaction is mediated mainly by van der Waals contacts, which is in agreement with the observed enthalpic contribution determined by non-linear chromatography. However, the calculated entropic contribution also indicates local conformational changes. Collectively our data suggest that ibogaine and PCP bind to overlapping sites located between the serine and valine/phenylalanine rings, to finally block the AChR ion channel, and in the case of ibogaine, to probably maintain the AChR in the desensitized state for longer time.
Figures
References
-
- Arias HR. Thermodynamics of nicotinic receptor interactions. In: Raffa RB, editor. Drug-receptor thermodynamics: introduction and applications. USA: John Wiley & Sons, Ltd; 2001. pp. 293–358.
-
- Arias HR. Ligand-gated ion channel receptor superfamilies. In: Arias HR, editor. Biological and biophysical aspects of ligand-gated ion channel receptor super-families. Kerala, India: Research Signpost; 2006. pp. 1–25. [Chapter 1]
-
- Arias HR. Is the inhibition of nicotinic acetylcholine receptors by bupropion involved in its clinical actions? Int J Biochem Cell Biol. 2009;41:2098–2108. - PubMed
-
- Arias HR, Bhumireddy P, Bouzat C. Molecular mechanisms and binding site locations for noncompetitive antagonists of nicotinic acetylcholine receptors. Int J Biochem Cell Biol. 2006a;38:1254–1276. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
