

Identification of zyklopen, a new member of the vertebrate multicopper ferroxidase family, and characterization in rodents and human cells
- PMID: 20685892
- PMCID: PMC2937573
- DOI: 10.3945/jn.109.117531
Identification of zyklopen, a new member of the vertebrate multicopper ferroxidase family, and characterization in rodents and human cells
Abstract
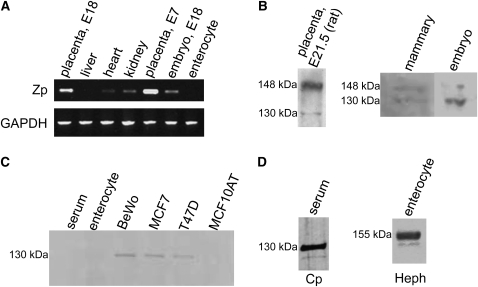
We previously detected a membrane-bound, copper-containing oxidase that may be involved in iron efflux in BeWo cells, a human placental cell line. We have now identified a gene encoding a predicted multicopper ferroxidase (MCF) with a putative C-terminal membrane-spanning sequence and high sequence identity to hephaestin (Heph) and ceruloplasmin (Cp), the other known vertebrate MCF. Molecular modeling revealed conservation of all type I, II, and III copper-binding sites as well as a putative iron-binding site. Protein expression was observed in multiple diverse mouse tissues, including placenta and mammary gland, and the expression pattern was distinct from that of Cp and Heph. The protein possessed ferroxidase activity, and protein levels decreased in cellular copper deficiency. Knockdown with small interfering RNA in BeWo cells indicates that this gene represents the previously detected oxidase. We propose calling this new member of the MCF family "zyklopen."
Conflict of interest statement
Author disclosures: H. Chen, Z. K. Attieh, B. A. Syed, Y-M. Kuo, V. Stevens, B. K. Fuqua, H. S. Andersen, C. E. Naylor, R. W. Evans, L. Gambling, R. Danzeisen, M. Bacouri-Haidar, J. Usta, C. D. Vulpe and H. J. McArdle, no conflicts of interest.
Figures





Similar articles
-
The biology of mammalian multi-copper ferroxidases.Biometals. 2023 Apr;36(2):263-281. doi: 10.1007/s10534-022-00370-z. Epub 2022 Feb 15. Biometals. 2023. PMID: 35167013 Free PMC article. Review.
-
Impact of maternal iron deficiency anaemia on the expression of the newly discovered multi-copper ferroxidase, Zyklopen, in term placentas.J Obstet Gynaecol. 2022 Jan;42(1):74-82. doi: 10.1080/01443615.2021.1882968. Epub 2021 May 2. J Obstet Gynaecol. 2022. Retraction in: J Obstet Gynaecol. 2024 Dec;44(1):2382547. doi: 10.1080/01443615.2024.2382547. PMID: 33938360 Retracted.
-
[Ceruloplasmin, hephaestin and zyklopen: the three multicopper oxidases important for human iron metabolism].Postepy Hig Med Dosw (Online). 2014;68:912-24. doi: 10.5604/17322693.1111136. Postepy Hig Med Dosw (Online). 2014. PMID: 24988611 Review. Polish.
-
Severe Iron Metabolism Defects in Mice With Double Knockout of the Multicopper Ferroxidases Hephaestin and Ceruloplasmin.Cell Mol Gastroenterol Hepatol. 2018 Jun 23;6(4):405-427. doi: 10.1016/j.jcmgh.2018.06.006. eCollection 2018. Cell Mol Gastroenterol Hepatol. 2018. PMID: 30182051 Free PMC article.
-
Recombinant expression and functional characterization of human hephaestin: a multicopper oxidase with ferroxidase activity.Biochemistry. 2005 Nov 15;44(45):14725-31. doi: 10.1021/bi051559k. Biochemistry. 2005. PMID: 16274220
Cited by
-
The biology of mammalian multi-copper ferroxidases.Biometals. 2023 Apr;36(2):263-281. doi: 10.1007/s10534-022-00370-z. Epub 2022 Feb 15. Biometals. 2023. PMID: 35167013 Free PMC article. Review.
-
Iron in the Tumor Microenvironment-Connecting the Dots.Front Oncol. 2018 Nov 26;8:549. doi: 10.3389/fonc.2018.00549. eCollection 2018. Front Oncol. 2018. PMID: 30534534 Free PMC article. Review.
-
From Rust to Quantum Biology: The Role of Iron in Retina Physiopathology.Cells. 2020 Mar 13;9(3):705. doi: 10.3390/cells9030705. Cells. 2020. PMID: 32183063 Free PMC article. Review.
-
Biomarkers of Nutrition for Development (BOND)-Iron Review.J Nutr. 2018 Jun 1;148(suppl_1):1001S-1067S. doi: 10.1093/jn/nxx036. J Nutr. 2018. PMID: 29878148 Free PMC article. Review.
-
Ceruloplasmin and hephaestin jointly protect the exocrine pancreas against oxidative damage by facilitating iron efflux.Redox Biol. 2018 Jul;17:432-439. doi: 10.1016/j.redox.2018.05.013. Epub 2018 May 31. Redox Biol. 2018. PMID: 29883959 Free PMC article.
References
-
- Kosman DJ. Fet3p, ceruloplasmin and the role of copper in iron metabolism. Adv Protein Chem. 2002;60:221–69 - PubMed
-
- Hellman NE, Gitlin JD. Ceruloplasmin metabolism and function. Annu Rev Nutr. 2002;22:439–58 - PubMed
-
- Vulpe CD, Kuo Y-M, Libina N, Askwith C, Murphy TL, Cowley L, Gitschier J, Anderson G. Hephaestin, a ceruloplasmin homologue implicated in intestinal iron transport, is defective in the sla mouse. Nat Genet. 1999;21:195–9 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
