

Persistent representation of juvenile experience in the adult songbird brain
- PMID: 20686001
- PMCID: PMC2927487
- DOI: 10.1523/JNEUROSCI.6042-09.2010
Persistent representation of juvenile experience in the adult songbird brain
Abstract
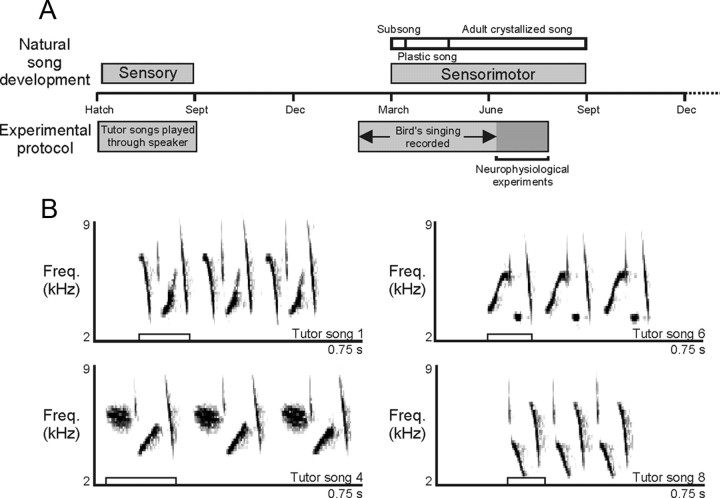
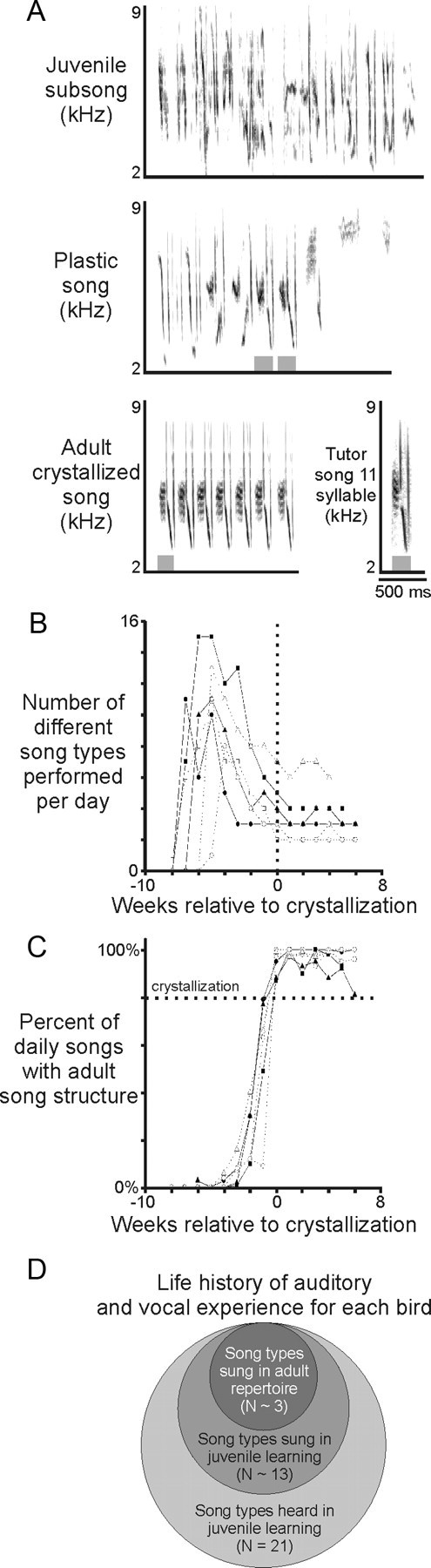
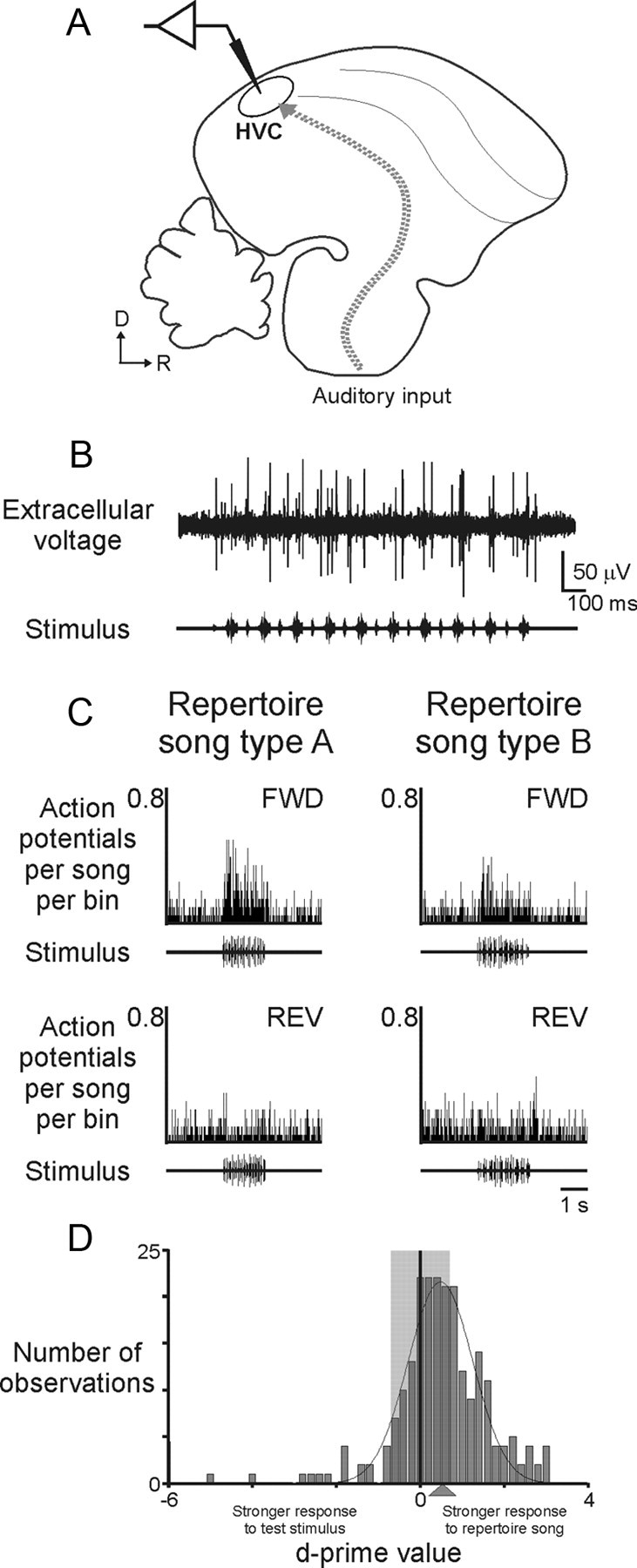
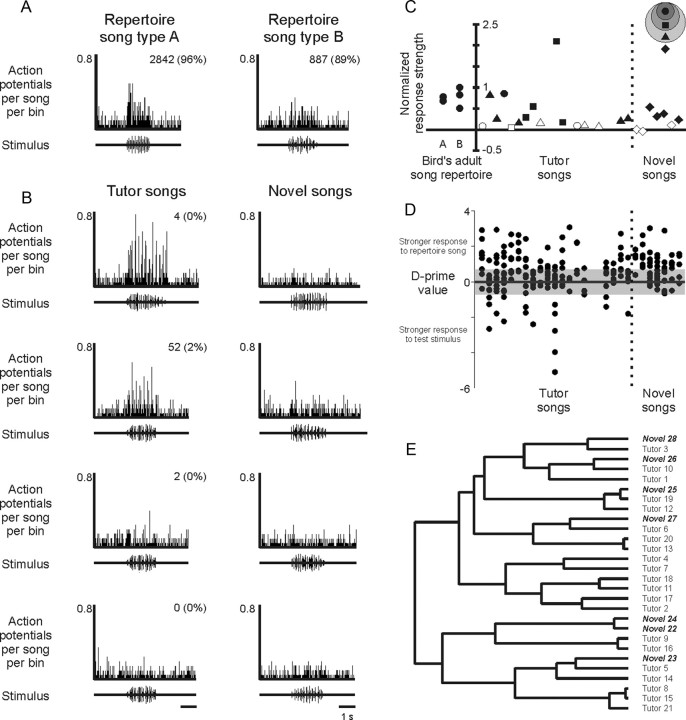
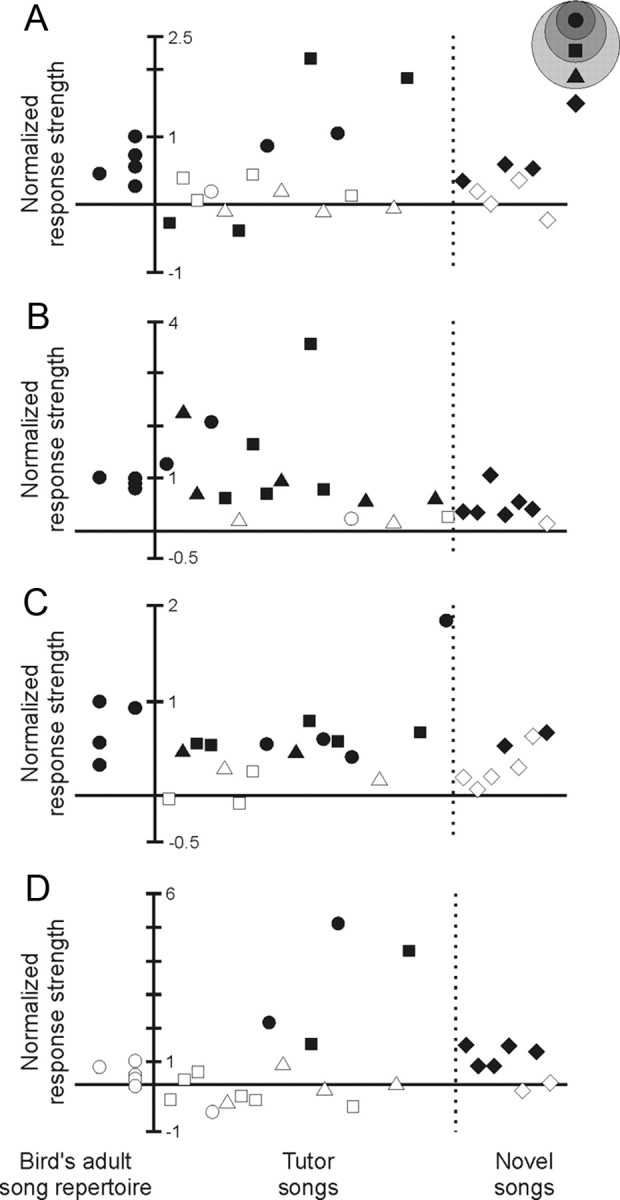
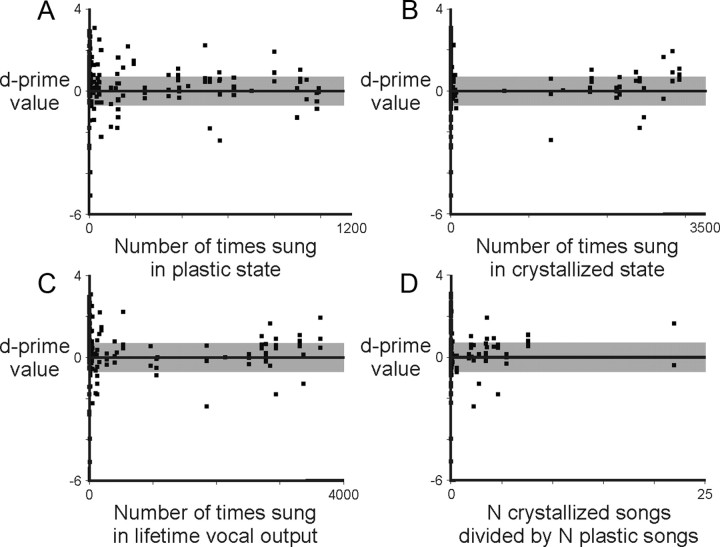
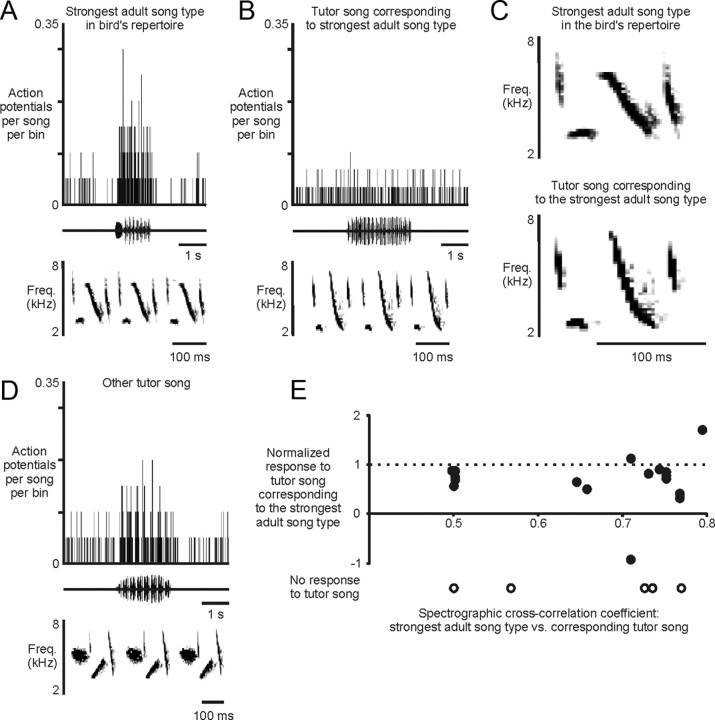
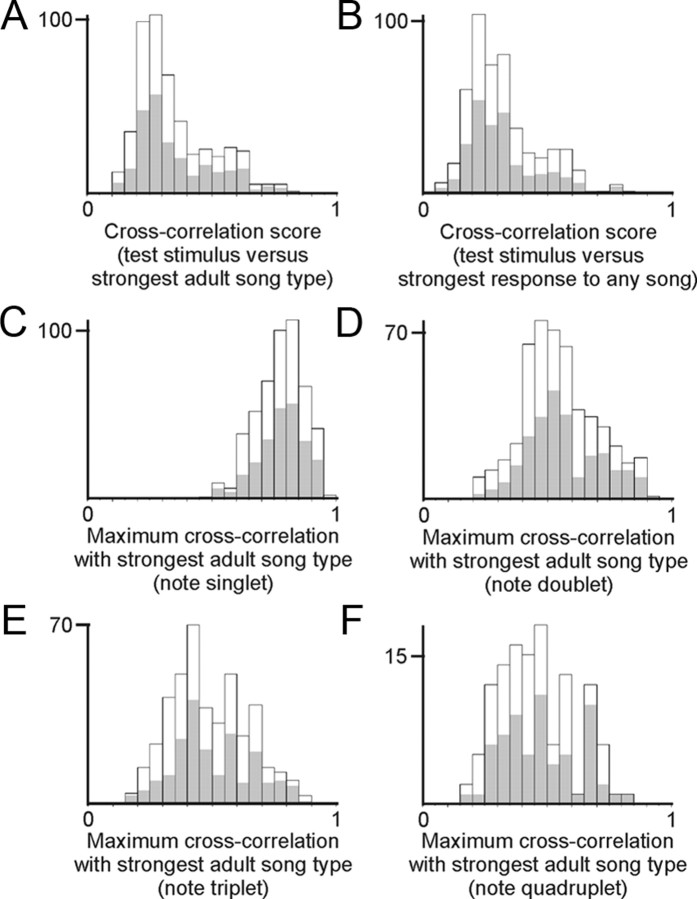
Juveniles sometimes learn behaviors that they cease to express as adults. Whether the adult brain retains a record of experiences associated with behaviors performed transiently during development remains unclear. We addressed this issue by studying neural representations of song in swamp sparrows, a species in which juveniles learn and practice many more songs than they retain in their adult vocal repertoire. We exposed juvenile swamp sparrows to a suite of tutor songs and confirmed that, although many tutor songs were imitated during development, not all copied songs were retained into adulthood. We then recorded extracellularly in the sensorimotor nucleus HVC in anesthetized sparrows to assess neuronal responsiveness to songs in the adult repertoire, tutor songs, and novel songs. Individual HVC neurons almost always responded to songs in the adult repertoire and commonly responded even more strongly to a tutor song. Effective tutor songs were not simply those that were acoustically similar to songs in the adult repertoire. Moreover, the strength of tutor song responses was unrelated to the number of times that the bird sang copies of those songs in juvenile or adult life. Notably, several neurons responded most strongly to a tutor song performed only rarely and transiently during juvenile life, or even to a tutor song for which we could find no evidence of ever having been copied. Thus, HVC neurons representing songs in the adult repertoire also appear to retain a lasting record of certain tutor songs, including those imitated only transiently.
Figures
Similar articles
-
Experience-Dependent Intrinsic Plasticity During Auditory Learning.J Neurosci. 2019 Feb 13;39(7):1206-1221. doi: 10.1523/JNEUROSCI.1036-18.2018. Epub 2018 Dec 12. J Neurosci. 2019. PMID: 30541908 Free PMC article.
-
Anterior forebrain neurons develop selectivity by an intermediate stage of birdsong learning.J Neurosci. 1997 Aug 15;17(16):6447-62. doi: 10.1523/JNEUROSCI.17-16-06447.1997. J Neurosci. 1997. PMID: 9236252 Free PMC article.
-
Distributed and selective auditory representation of song repertoires in the avian song system.J Neurophysiol. 2006 Dec;96(6):3433-47. doi: 10.1152/jn.01130.2005. Epub 2006 Aug 2. J Neurophysiol. 2006. PMID: 16885516
-
Tutor auditory memory for guiding sensorimotor learning in birdsong.Front Neural Circuits. 2024 Jul 1;18:1431119. doi: 10.3389/fncir.2024.1431119. eCollection 2024. Front Neural Circuits. 2024. PMID: 39011279 Free PMC article. Review.
-
Song- and order-selective neurons develop in the songbird anterior forebrain during vocal learning.J Neurobiol. 1997 Nov;33(5):694-709. J Neurobiol. 1997. PMID: 9369467 Review.
Cited by
-
Memory circuits for vocal imitation.Curr Opin Neurobiol. 2020 Feb;60:37-46. doi: 10.1016/j.conb.2019.11.002. Epub 2019 Dec 4. Curr Opin Neurobiol. 2020. PMID: 31810009 Free PMC article. Review.
-
Neuroestrogen synthesis modifies neural representations of learned song without altering vocal imitation in developing songbirds.Sci Rep. 2020 Feb 27;10(1):3602. doi: 10.1038/s41598-020-60329-3. Sci Rep. 2020. PMID: 32108169 Free PMC article.
-
Accelerated redevelopment of vocal skills is preceded by lasting reorganization of the song motor circuitry.Elife. 2019 May 17;8:e43194. doi: 10.7554/eLife.43194. Elife. 2019. PMID: 31099755 Free PMC article.
-
Insights into the Neural and Genetic Basis of Vocal Communication.Cell. 2016 Mar 10;164(6):1269-1276. doi: 10.1016/j.cell.2016.02.039. Cell. 2016. PMID: 26967292 Free PMC article. Review.
-
Motor circuits are required to encode a sensory model for imitative learning.Nat Neurosci. 2012 Oct;15(10):1454-9. doi: 10.1038/nn.3206. Epub 2012 Sep 16. Nat Neurosci. 2012. PMID: 22983208 Free PMC article.
References
-
- Balaban E. Cultural and genetic variation in swamp sparrows (Melospiza georgiana). 1. Song variation, genetic variation, and their relationship. Behaviour. 1988a;105:250–291.
-
- Balaban E. Cultural and genetic variation in swamp sparrows (Melospiza georgiana). 2. Behavioral salience of geographic song variants. Behaviour. 1988b;105:292–322.
-
- Ballentine B, Hyman J, Nowicki S. Vocal performance influences female response to male bird song: an experimental test. Behav Ecol. 2004;15:163–168.
-
- Bolhuis JJ, Gahr M. Neural mechanisms of birdsong memory. Nat Rev Neurosci. 2006;7:347–357. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources