

Nuclear localization of tegument-delivered pp71 in human cytomegalovirus-infected cells is facilitated by one or more factors present in terminally differentiated fibroblasts
- PMID: 20686028
- PMCID: PMC2937784
- DOI: 10.1128/JVI.00500-10
Nuclear localization of tegument-delivered pp71 in human cytomegalovirus-infected cells is facilitated by one or more factors present in terminally differentiated fibroblasts
Abstract
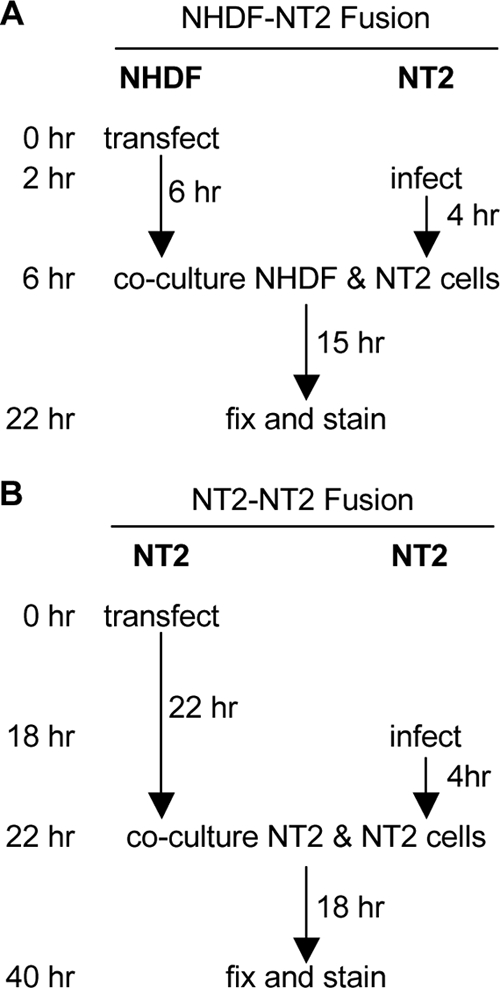
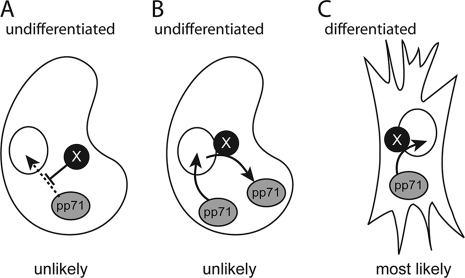
Herpesviral virions contain a tegument layer that consists primarily of viral proteins. The delivery of fully functional proteins to infected cells upon virion envelope fusion to the plasma membrane allows herpesviruses to modulate cellular activities prior to viral gene expression. Certain tegument proteins can also regulate viral processes. For example, the pp71 tegument protein encoded by the UL82 gene of human cytomegalovirus (HCMV) stimulates viral immediate early (IE) gene expression and thus acts to initiate the productive lytic infectious cycle. In terminally differentiated fibroblasts infected with HCMV, tegument-delivered pp71 traffics to the nucleus and degrades the cellular transcriptional corepressor Daxx to initiate viral IE gene expression and lytic replication. However, when HCMV infects incompletely differentiated cells, tegument-delivered pp71 remains in the cytoplasm, allowing the nucleus-localized Daxx protein to silence viral IE gene expression and promote the establishment of a latent infection in certain cell types. We sought to determine whether undifferentiated cells block the trafficking of tegument-delivered pp71 to the nucleus or whether differentiated cells facilitate the nuclear transport of tegument-delivered pp71. Heterogenous cell fusion experiments demonstrated that tegument-delivered pp71 found in the cytoplasm of undifferentiated NT2 cells could be driven into the nucleus by one or more factors provided by fully differentiated fibroblasts. Our data raise the intriguing possibility that latency is the default program launched by HCMV upon viral entry into cells and that lytic infection is initiated only in certain (differentiated) cells that can facilitate the delivery of incoming pp71 to the nucleus.
Figures







Similar articles
-
Daxx-mediated accumulation of human cytomegalovirus tegument protein pp71 at ND10 facilitates initiation of viral infection at these nuclear domains.J Virol. 2002 Aug;76(15):7705-12. doi: 10.1128/jvi.76.15.7705-7712.2002. J Virol. 2002. PMID: 12097584 Free PMC article.
-
Human cytomegalovirus gene expression is silenced by Daxx-mediated intrinsic immune defense in model latent infections established in vitro.J Virol. 2007 Sep;81(17):9109-20. doi: 10.1128/JVI.00827-07. Epub 2007 Jun 27. J Virol. 2007. PMID: 17596307 Free PMC article.
-
Cellular and viral control over the initial events of human cytomegalovirus experimental latency in CD34+ cells.J Virol. 2010 Jun;84(11):5594-604. doi: 10.1128/JVI.00348-10. Epub 2010 Mar 24. J Virol. 2010. PMID: 20335255 Free PMC article.
-
Expanding the Known Functional Repertoire of the Human Cytomegalovirus pp71 Protein.Front Cell Infect Microbiol. 2020 Mar 12;10:95. doi: 10.3389/fcimb.2020.00095. eCollection 2020. Front Cell Infect Microbiol. 2020. PMID: 32226778 Free PMC article. Review.
-
Human cytomegalovirus tegument proteins (pp65, pp71, pp150, pp28).Virol J. 2012 Jan 17;9:22. doi: 10.1186/1743-422X-9-22. Virol J. 2012. PMID: 22251420 Free PMC article. Review.
Cited by
-
Human cytomegalovirus infection and colorectal cancer risk: a meta-analysis.Oncotarget. 2016 Nov 22;7(47):76735-76742. doi: 10.18632/oncotarget.12523. Oncotarget. 2016. PMID: 27732934 Free PMC article.
-
Early inhibitors of human cytomegalovirus: state-of-art and therapeutic perspectives.Pharmacol Ther. 2011 Sep;131(3):309-29. doi: 10.1016/j.pharmthera.2011.04.007. Epub 2011 Apr 28. Pharmacol Ther. 2011. PMID: 21570424 Free PMC article. Review.
-
Regulation of nucleocytoplasmic trafficking of viral proteins: an integral role in pathogenesis?Biochim Biophys Acta. 2011 Dec;1813(12):2176-90. doi: 10.1016/j.bbamcr.2011.03.019. Epub 2011 Apr 16. Biochim Biophys Acta. 2011. PMID: 21530593 Free PMC article. Review.
-
Noncytotoxic functions of killer cell granzymes in viral infections.PLoS Pathog. 2021 Sep 16;17(9):e1009818. doi: 10.1371/journal.ppat.1009818. eCollection 2021 Sep. PLoS Pathog. 2021. PMID: 34529743 Free PMC article. Review.
-
Tumor Necrosis Factor Alpha Induces Reactivation of Human Cytomegalovirus Independently of Myeloid Cell Differentiation following Posttranscriptional Establishment of Latency.mBio. 2018 Sep 11;9(5):e01560-18. doi: 10.1128/mBio.01560-18. mBio. 2018. PMID: 30206173 Free PMC article.
References
-
- Ahn, J. H., and G. S. Hayward. 2000. Disruption of PML-associated nuclear bodies by IE1 correlates with efficient early stages of viral gene expression and DNA replication in human cytomegalovirus infection. Virology 274:39-55. - PubMed
-
- Bernardi, R., and P. P. Pandolfi. 2007. Structure, dynamics and functions of promyelocytic leukaemia nuclear bodies. Nat. Rev. Mol. Cell Biol. 8:1006-1016. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
