

Adenovirus-based vaccines: comparison of vectors from three species of adenoviridae
- PMID: 20686035
- PMCID: PMC2950567
- DOI: 10.1128/JVI.00450-10
Adenovirus-based vaccines: comparison of vectors from three species of adenoviridae
Abstract
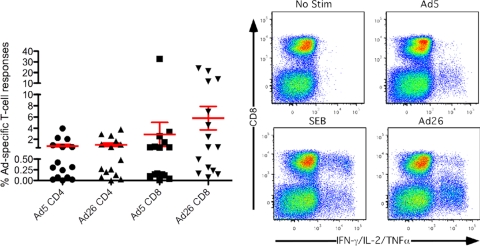
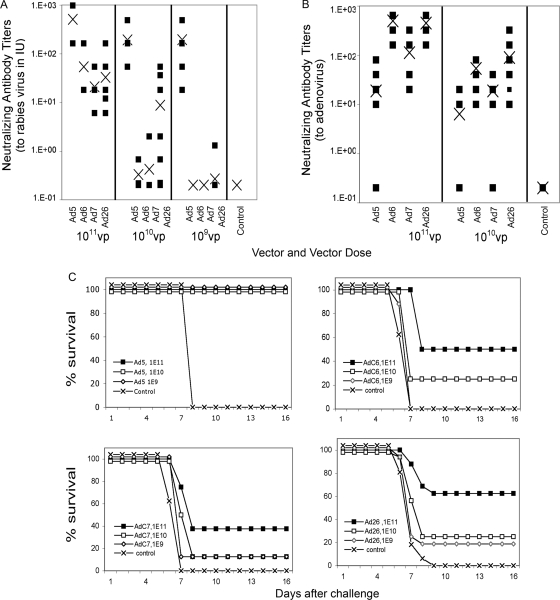
In order to better understand the broad applicability of adenovirus (Ad) as a vector for human vaccine studies, we compared four adenovirus (Ad) vectors from families C (Ad human serotype 5 [HAdV-5; here referred to as AdHu5]), D (HAdV-26; here referred to as AdHu26), and E (simian serotypes SAdV-23 and SAdV-24; here referred to as chimpanzee serotypes 6 and 7 [AdC6 and AdC7, respectively]) of the Adenoviridae. Seroprevalence rates and titers of neutralizing antibodies to the two human-origin Ads were found to be higher than those reported previously, especially in countries of sub-Saharan Africa. Conversely, prevalence rates and titers to AdC6 and AdC7 were markedly lower. Healthy human adults from the United States had readily detectable circulating T cells recognizing Ad viruses, the levels of which in some individuals were unexpectedly high in response to AdHu26. The magnitude of T-cell responses to AdHu5 correlated with those to AdHu26, suggesting T-cell recognition of conserved epitopes. In mice, all of the different Ad vectors induced CD8(+) T-cell responses that were comparable in their magnitudes and cytokine production profiles. Prime-boost regimens comparing different combinations of Ad vectors failed to indicate that the sequential use of Ad vectors from distinct families resulted in higher immune responses than the use of serologically distinct Ad vectors from the same family. Moreover, the transgene product-specific antibody responses induced by the AdHu26 and AdC vectors were markedly lower than those induced by the AdHu5 vector. AdHu26 vectors and, to a lesser extent, AdC vectors induced more potent Ad-neutralizing antibody responses. These results suggest that the potential of AdHu26 as a vaccine vector may suffer from limitations similar to those found for vectors based on other prevalent human Ads.
Figures





References
-
- Abbink, P., A. A. Lemckert, B. A. Ewald, D. M. Lynch, M. Denholtz, S. Smits, L. Holterman, I. Damen, R. Vogels, A. R. Thorner, K. L. O'Brien, A. Carville, K. G. Mansfield, J. Goudsmit, M. J. Havenga, and D. H. Barouch. 2007. Comparative seroprevalence and immunogenicity of six rare serotype recombinant adenovirus vaccine vectors from subgroups B and D. J. Virol. 81:4654-4663. - PMC - PubMed
-
- Amara, R. R., J. M. Smith, S. I. Staprans, D. C. Montefiori, F. Villinger, J. D. Altman, S. P. O'Neil, N. L. Kozyr, Y. Xu, L. S. Wyatt, P. L. Earl, J. G. Herndon, J. M. McNicholl, H. M. McClure, B. Moss, and H. L. Robinson. 2002. Critical role for Env as well as Gag-Pol in control of a simian-human immunodeficiency virus 89.6P challenge by a DNA prime/recombinant modified vaccinia virus Ankara vaccine. J. Virol. 76:6138-6146. - PMC - PubMed
-
- Bansal, G. P., A. Malaspina, and J. Flores. Future paths for HIV vaccine research: exploiting results from recent clinical trials and current scientific advances. Curr. Opin. Mol. Ther. 12:39-46. - PubMed
-
- Buchbinder, S. P., D. V. Mehrotra, A. Duerr, D. W. Fitzgerald, R. Mogg, D. Li, P. B. Gilbert, J. R. Lama, M. Marmor, C. Del Rio, M. J. McElrath, D. R. Casimiro, K. M. Gottesdiener, J. A. Chodakewitz, L. Corey, and M. N. Robertson. 2008. Efficacy assessment of a cell-mediated immunity HIV-1 vaccine (the Step Study): a double-blind, randomised, placebo-controlled, test-of-concept trial. Lancet 372:1881-1893. - PMC - PubMed
-
- Fitzgerald, J. C., G. P. Gao, A. Reyes-Sandoval, G. N. Pavlakis, Z. Q. Xiang, A. P. Wlazlo, W. Giles-Davis, J. M. Wilson, and H. C. Ertl. 2003. A simian replication-defective adenoviral recombinant vaccine to HIV-1 gag. J. Immunol. 170:1416-1422. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
