

Mutations of Francisella novicida that alter the mechanism of its phagocytosis by murine macrophages
- PMID: 20686600
- PMCID: PMC2912274
- DOI: 10.1371/journal.pone.0011857
Mutations of Francisella novicida that alter the mechanism of its phagocytosis by murine macrophages
Abstract
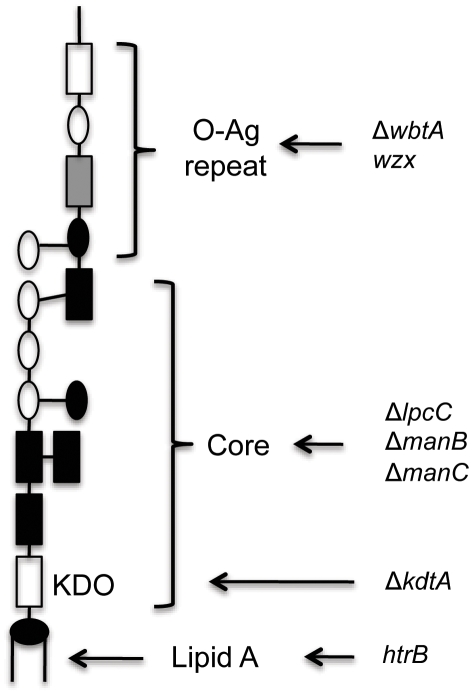
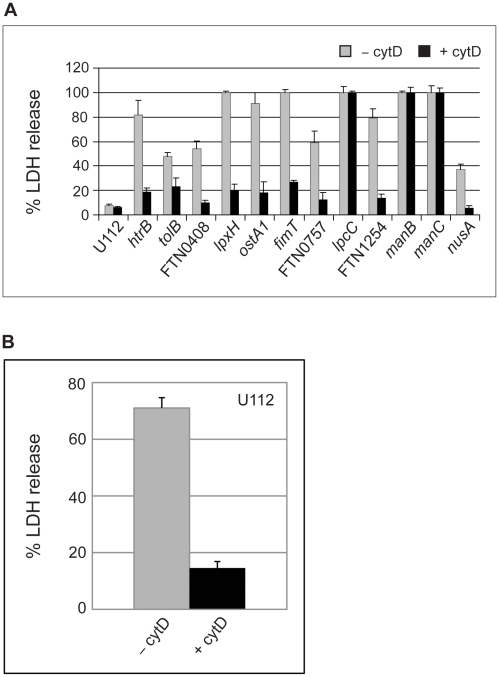
Infection with the bacterial pathogen Francisella tularensis tularensis (F. tularensis) causes tularemia, a serious and debilitating disease. Francisella tularensis novicida strain U112 (abbreviated F. novicida), which is closely related to F. tularensis, is pathogenic for mice but not for man, making it an ideal model system for tularemia. Intracellular pathogens like Francisella inhibit the innate immune response, thereby avoiding immune recognition and death of the infected cell. Because activation of inflammatory pathways may lead to cell death, we reasoned that we could identify bacterial genes involved in inhibiting inflammation by isolating mutants that killed infected cells faster than the wild-type parent. We screened a comprehensive transposon library of F. novicida for mutant strains that increased the rate of cell death following infection in J774 macrophage-like cells, as compared to wild-type F. novicida. Mutations in 28 genes were identified as being hypercytotoxic to both J774 and primary macrophages of which 12 were less virulent in a mouse infection model. Surprisingly, we found that F. novicida with mutations in four genes (lpcC, manB, manC and kdtA) were taken up by and killed macrophages at a much higher rate than the parent strain, even upon treatment with cytochalasin D (cytD), a classic inhibitor of macrophage phagocytosis. At least 10-fold more mutant bacteria were internalized by macrophages as compared to the parent strain if the bacteria were first fixed with formaldehyde, suggesting a surface structure is required for the high phagocytosis rate. However, bacteria were required to be viable for macrophage toxicity. The four mutant strains do not make a complete LPS but instead have an exposed lipid A. Interestingly, other mutations that result in an exposed LPS core were not taken up at increased frequency nor did they kill host cells more than the parent. These results suggest an alternative, more efficient macrophage uptake mechanism for Francisella that requires exposure of a specific bacterial surface structure(s) but results in increased cell death following internalization of live bacteria.
Conflict of interest statement
Figures
Similar articles
-
Comparative phosphoproteomics reveals components of host cell invasion and post-transcriptional regulation during Francisella infection.Mol Cell Proteomics. 2013 Nov;12(11):3297-309. doi: 10.1074/mcp.M113.029850. Epub 2013 Aug 22. Mol Cell Proteomics. 2013. PMID: 23970565 Free PMC article.
-
TolC and EmrA1 contribute to Francisella novicida multidrug resistance and modulation of host cell death.J Bacteriol. 2024 Sep 19;206(9):e0024624. doi: 10.1128/jb.00246-24. Epub 2024 Aug 28. J Bacteriol. 2024. PMID: 39194223 Free PMC article.
-
Francisella novicida LPS has greater immunobiological activity in mice than F. tularensis LPS, and contributes to F. novicida murine pathogenesis.Microbes Infect. 2003 Apr;5(5):397-403. doi: 10.1016/s1286-4579(03)00052-2. Microbes Infect. 2003. PMID: 12737995
-
Comparative review of Francisella tularensis and Francisella novicida.Front Cell Infect Microbiol. 2014 Mar 13;4:35. doi: 10.3389/fcimb.2014.00035. eCollection 2014. Front Cell Infect Microbiol. 2014. PMID: 24660164 Free PMC article. Review.
-
Monocyte/macrophage inflammatory response pathways to combat Francisella infection: possible therapeutic targets?Front Cell Infect Microbiol. 2014 Feb 21;4:18. doi: 10.3389/fcimb.2014.00018. eCollection 2014. Front Cell Infect Microbiol. 2014. PMID: 24600590 Free PMC article. Review.
Cited by
-
Customizable PCR-microplate array for differential identification of multiple pathogens.J Food Prot. 2013 Nov;76(11):1948-57. doi: 10.4315/0362-028X.JFP-13-153. J Food Prot. 2013. PMID: 24215700 Free PMC article.
-
Microinjection of Francisella tularensis and Listeria monocytogenes reveals the importance of bacterial and host factors for successful replication.Infect Immun. 2015 Aug;83(8):3233-42. doi: 10.1128/IAI.00416-15. Epub 2015 Jun 1. Infect Immun. 2015. PMID: 26034213 Free PMC article.
-
The Lipid A 1-Phosphatase, LpxE, Functionally Connects Multiple Layers of Bacterial Envelope Biogenesis.mBio. 2019 Jun 18;10(3):e00886-19. doi: 10.1128/mBio.00886-19. mBio. 2019. PMID: 31213552 Free PMC article.
-
Impact of Francisella tularensis pilin homologs on pilus formation and virulence.Microb Pathog. 2011 Sep;51(3):110-20. doi: 10.1016/j.micpath.2011.05.001. Epub 2011 May 13. Microb Pathog. 2011. PMID: 21605655 Free PMC article.
-
The Francisella O-antigen mediates survival in the macrophage cytosol via autophagy avoidance.Cell Microbiol. 2014 Jun;16(6):862-77. doi: 10.1111/cmi.12246. Epub 2013 Dec 16. Cell Microbiol. 2014. PMID: 24286610 Free PMC article.
References
-
- Sjostedt A. Family III. Francisellaceae. In: Brenner DJ, Krieg NR, Staley JT, Garrity GM, editors. Bergey's Manual of Systematic Bacteriology, 2nd ed, vol. 2 (The Proteobacteria), part B (The Gammaproteobacteria) New York: Springer; 2005. pp. 199–209.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
