

Ligand-based peptide design and combinatorial peptide libraries to target G protein-coupled receptors
- PMID: 20687879
- PMCID: PMC4939874
- DOI: 10.2174/138161210793292474
Ligand-based peptide design and combinatorial peptide libraries to target G protein-coupled receptors
Abstract
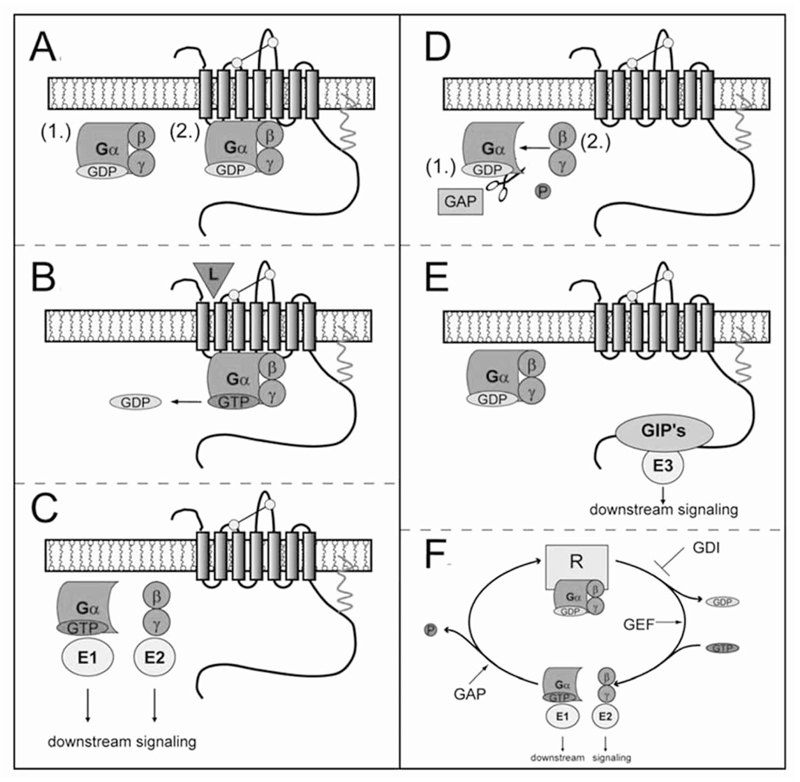
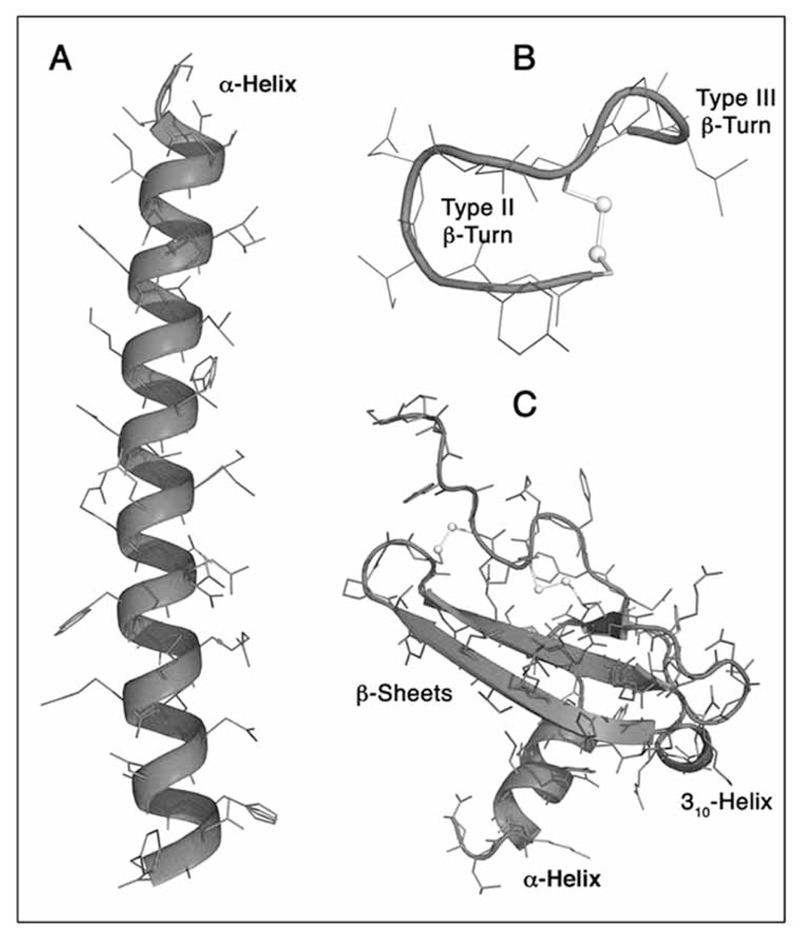
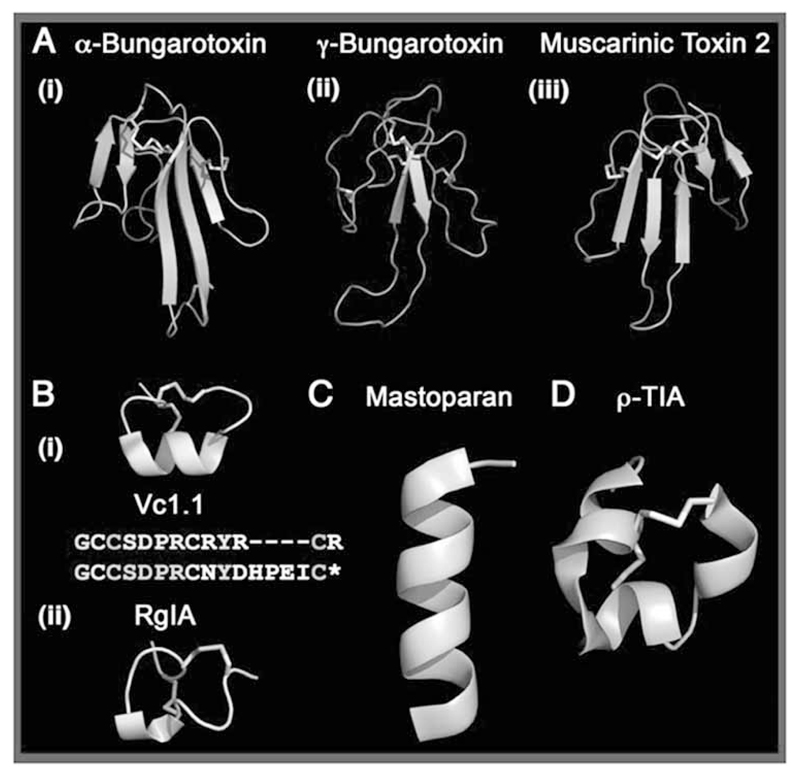
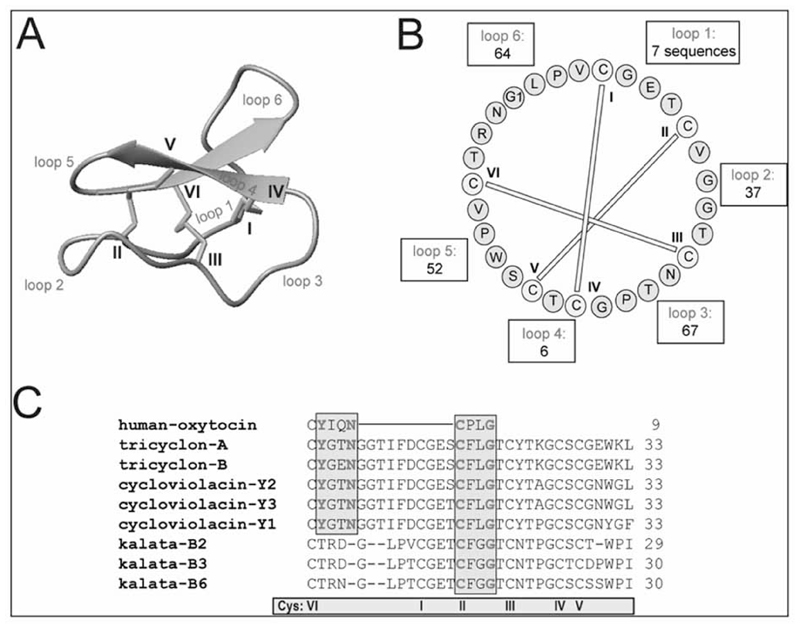
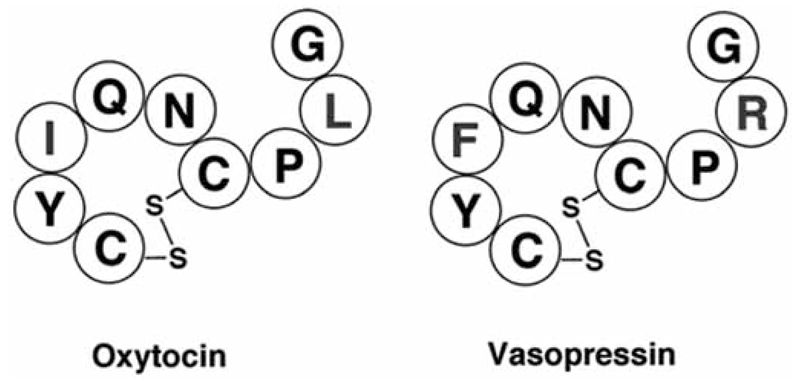
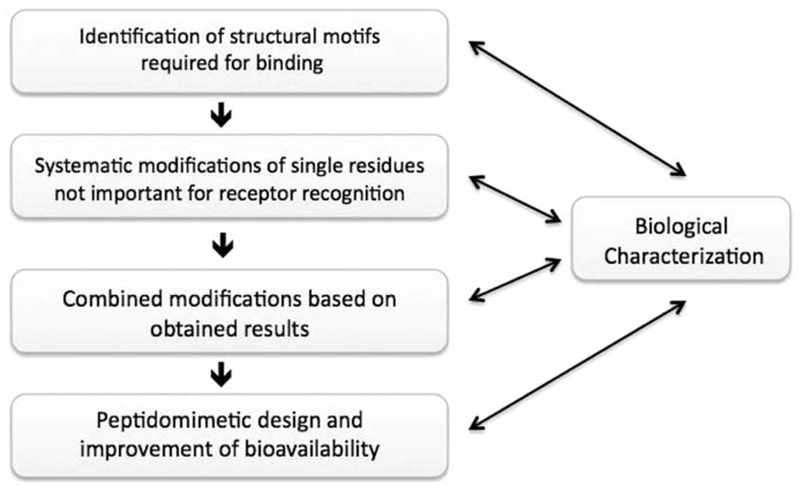
G protein-coupled receptors (GPCRs) are considered to represent the most promising drug targets; it has been repeatedly said that a large fraction of the currently marketed drugs elicit their actions by binding to GPCRs (with cited numbers varying from 30-50%). Closer scrutiny, however, shows that only a modest fraction of (≈60) GPCRs are, in fact, exploited as drug targets, only ≈20 of which are peptide-binding receptors. The vast majority of receptors in the humane genome have not yet been explored as sites of action for drugs. Given the drugability of this receptor class, it appears that opportunities for drug discovery abound. In addition, GPCRs provide for binding sites other than the ligand binding sites (referred to as the "orthosteric site"). These additional sites include (i) binding sites for ligands (referred to as "allosteric ligands") that modulate the affinity and efficacy of orthosteric ligands, (ii) the interaction surface that recruits G proteins and arrestins, (iii) the interaction sites of additional proteins (GIPs, GPCR interacting proteins that regulate G protein signaling or give rise to G protein-independent signals). These sites can also be targeted by peptides. Combinatorial and natural peptide libraries are therefore likely to play a major role in identifying new GPCR ligands at each of these sites. In particular the diverse natural peptide libraries such as the venom peptides from marine cone-snails and plant cyclotides have been established as a rich source of drug leads. High-throughput screening and combinatorial chemistry approaches allow for progressing from these starting points to potential drug candidates. This will be illustrated by focusing on the ligand-based drug design of oxytocin (OT) and vasopressin (AVP) receptor ligands using natural peptide leads as starting points.
Figures
References
-
- Kroeze WK, Sheffler DJ, Roth BL. G-protein-coupled receptors at a glance. J Cell Sci. 2003;116:4867–9. - PubMed
-
- Oldham WM, Hamm HE. Heterotrimeric G protein activation by G-protein-coupled receptors. Nat Rev Mol Cell Biol. 2008;9:60–71. - PubMed
-
- Perez DM. From plants to man: the GPCR “tree of life”. Mol Pharmacol. 2005;67:1383–4. - PubMed
-
- Marinissen MJ, Gutkind JS. G-protein-coupled receptors and signaling networks: emerging paradigms. Trends Pharmacol Sci. 2001;22:368–76. - PubMed
-
- Bjarnadottir TK, Gloriam DE, Hellstrand SH, et al. Comprehensive repertoire and phylogenetic analysis of the G protein-coupled receptors in human and mouse. Genomics. 2006;88:263–73. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous