

MiR-145, a new regulator of the DNA fragmentation factor-45 (DFF45)-mediated apoptotic network
- PMID: 20687965
- PMCID: PMC2924312
- DOI: 10.1186/1476-4598-9-211
MiR-145, a new regulator of the DNA fragmentation factor-45 (DFF45)-mediated apoptotic network
Abstract
Background: MicroRNA-145 (miR-145) is considered to play key roles in many cellular processes, such as proliferation, differentiation and apoptosis, by inhibiting target gene expression. DNA Fragmentation Factor-45 (DFF45) has been found to be the substrate of Caspase-3, and the cleavage of DFF45 by caspase-3 during apoptosis releases DFF40 that degrades chromosomal DNA into nucleosomal fragments. There are currently no in-depth studies on the relationship between miR-145 and the DFF45 gene.
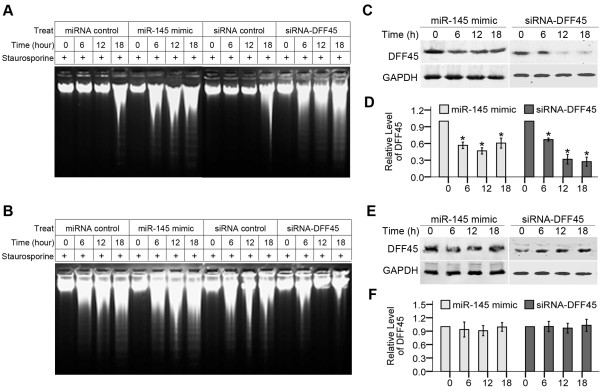
Results: In this study, we identified DFF45 as a novel target of miR-145. We demonstrated that miR-145 targets a putative binding site in the coding sequence (CDS) of DFF45, and its abundance is inversely associated with DFF45 expression in colon cancer cells. Using a luciferase reporter system, we found that miR-145 suppresses the expression of the luciferase reporter gene fused to the putative binding site of DFF45. The level of DFF45 protein, but not DFF45 mRNA, was decreased by miR-145, suggesting a mechanism of translational regulation. Furthermore, we demonstrate that this specific silencing of DFF45 by miR-145 accounts, at least in part, for the staurosporine-induced tumor cell apoptosis in vitro.
Conclusions: Our study reveals a previously unrecognized function of miR-145 in DFF45 processing, which may underlie crucial aspects of cancer biology.
Figures





Similar articles
-
Non-endometrioid and high-grade endometrioid endometrial cancers show DNA fragmentation factor 40 (DFF40) and B-cell lymphoma 2 protein (BCL2) underexpression, which predicts disease-free and overall survival, but not DNA fragmentation factor 45 (DFF45) underexpression.BMC Cancer. 2018 Apr 13;18(1):418. doi: 10.1186/s12885-018-4333-6. BMC Cancer. 2018. PMID: 29653556 Free PMC article.
-
Functional interaction of DFF35 and DFF45 with caspase-activated DNA fragmentation nuclease DFF40.J Biol Chem. 1999 Jul 23;274(30):20759-62. doi: 10.1074/jbc.274.30.20759. J Biol Chem. 1999. PMID: 10409614
-
Roles of DNA fragmentation factor and poly(ADP-ribose) polymerase in an amplification phase of tumor necrosis factor-induced apoptosis.J Biol Chem. 2001 Oct 12;276(41):38185-92. doi: 10.1074/jbc.M100629200. Epub 2001 Jul 18. J Biol Chem. 2001. PMID: 11461900
-
MicroRNA and cancer--focus on apoptosis.J Cell Mol Med. 2009 Jan;13(1):12-23. doi: 10.1111/j.1582-4934.2008.00510.x. J Cell Mol Med. 2009. PMID: 19175697 Free PMC article. Review.
-
The DFF40/CAD endonuclease and its role in apoptosis.Acta Biochim Pol. 2000;47(4):1037-44. Acta Biochim Pol. 2000. PMID: 11996094 Review.
Cited by
-
A Novel Tumor Suppressor SPINK5 Serves as an Independent Prognostic Predictor for Patients with Head and Neck Squamous Cell Carcinoma.Cancer Manag Res. 2020 Jun 23;12:4855-4869. doi: 10.2147/CMAR.S236266. eCollection 2020. Cancer Manag Res. 2020. PMID: 32606974 Free PMC article.
-
Double-negative feedback interaction between DNA methyltransferase 3A and microRNA-145 in the Warburg effect of ovarian cancer cells.Cancer Sci. 2018 Sep;109(9):2734-2745. doi: 10.1111/cas.13734. Epub 2018 Aug 12. Cancer Sci. 2018. PMID: 29993160 Free PMC article.
-
MicroRNAs and hypospadias: A systematic review.Med Int (Lond). 2024 Nov 18;5(1):7. doi: 10.3892/mi.2024.206. eCollection 2025 Jan-Feb. Med Int (Lond). 2024. PMID: 39624466 Free PMC article.
-
The effects of lidocaine and procaine on microRNA expression of adipocyte-derived adult stem cells.Korean J Anesthesiol. 2012 Jun;62(6):552-7. doi: 10.4097/kjae.2012.62.6.552. Epub 2012 Jun 19. Korean J Anesthesiol. 2012. PMID: 22778892 Free PMC article.
-
Network Pharmacology-Based Validation of Caveolin-1 as a Key Mediator of Ai Du Qing Inhibition of Drug Resistance in Breast Cancer.Front Pharmacol. 2018 Oct 2;9:1106. doi: 10.3389/fphar.2018.01106. eCollection 2018. Front Pharmacol. 2018. PMID: 30333750 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
