

Using automated imaging to interrogate gonadotrophin-releasing hormone receptor trafficking and function
- PMID: 20688134
- PMCID: PMC3021717
- DOI: 10.1016/j.mce.2010.07.008
Using automated imaging to interrogate gonadotrophin-releasing hormone receptor trafficking and function
Abstract
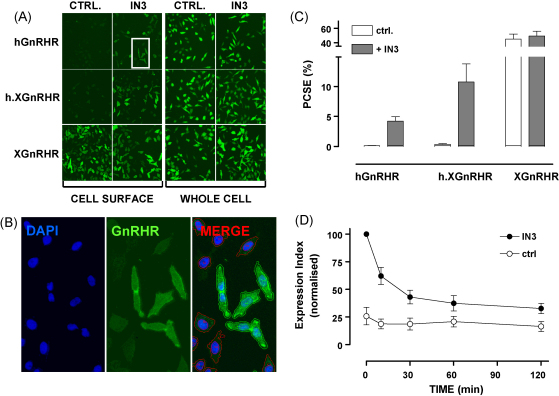
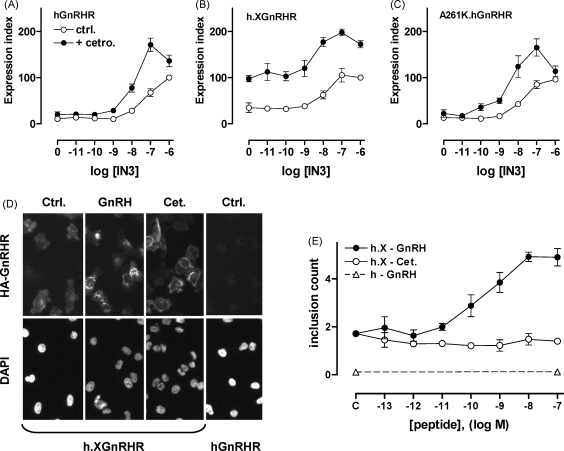
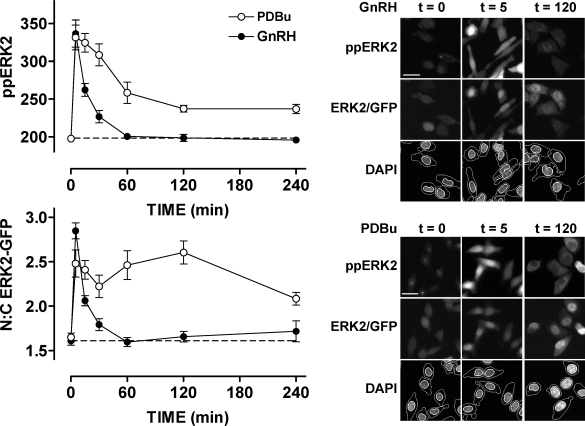
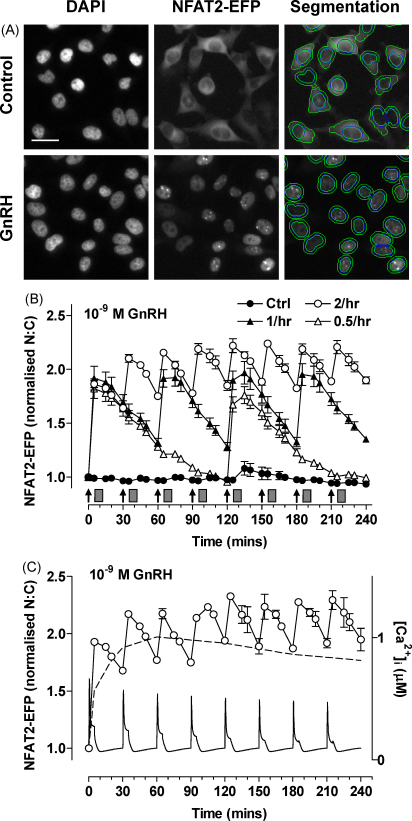
Gonadotrophin-releasing hormone (GnRH) acts via seven transmembrane receptors on gonadotrophs to stimulate gonadotrophin synthesis and secretion, and thereby mediates central control of reproduction. Type I mammalian GnRHR are unique, in that they lack C-terminal tails. This is thought to underlie their resistance to rapid homologous desensitisation as well as their slow rate of internalisation and inability to provoke G-protein-independent (arrestin-mediated) signalling. More recently it has been discovered that the vast majority of human GnRHR are actually intracellular, in spite of the fact that they are activated at the cell surface by a membrane impermeant peptide hormone. This apparently reflects inefficient exit from the endoplasmic reticulum and again, the absence of the C-tail likely contributes to their intracellular localisation. This review is intended to cover some of these novel aspects of GnRHR biology, focusing on ways that we have used automated fluorescence microscopy (high content imaging) to explore GnRHR localisation and trafficking as well as spatial and temporal aspects of GnRH signalling via the Ca(2+)/calmodulin/calcineurin/NFAT and Raf/MEK/ERK pathways.
Copyright © 2010 Elsevier Ireland Ltd. All rights reserved.
Figures
Similar articles
-
Trafficking and signalling of gonadotrophin-releasing hormone receptors: an automated imaging approach.Br J Pharmacol. 2010 Feb;159(4):751-60. doi: 10.1111/j.1476-5381.2009.00413.x. Epub 2009 Nov 3. Br J Pharmacol. 2010. PMID: 19888967 Free PMC article. Review.
-
Signalling, cycling and desensitisation of gonadotrophin-releasing hormone receptors.J Endocrinol. 2002 Apr;173(1):1-11. doi: 10.1677/joe.0.1730001. J Endocrinol. 2002. PMID: 11927379 Review.
-
Plasma membrane expression of gonadotropin-releasing hormone receptors: regulation by peptide and nonpeptide antagonists.Mol Endocrinol. 2010 Feb;24(2):423-35. doi: 10.1210/me.2009-0343. Epub 2009 Dec 15. Mol Endocrinol. 2010. PMID: 20009083 Free PMC article.
-
The gonadotrophin-releasing hormone receptor: signalling, cycling and desensitisation.Arch Physiol Biochem. 2002 Apr;110(1-2):113-22. doi: 10.1076/apab.110.1.113.893. Arch Physiol Biochem. 2002. PMID: 11935408 Review.
-
Agonist-induced internalization and downregulation of gonadotropin-releasing hormone receptors.Am J Physiol Cell Physiol. 2009 Sep;297(3):C591-600. doi: 10.1152/ajpcell.00166.2009. Epub 2009 Jul 8. Am J Physiol Cell Physiol. 2009. PMID: 19587220 Free PMC article.
Cited by
-
GATA2-induced silencing and LIM-homeodomain protein-induced activation are mediated by a bi-functional response element in the rat GnRH receptor gene.Mol Endocrinol. 2013 Jan;27(1):74-91. doi: 10.1210/me.2012-1182. Epub 2012 Dec 4. Mol Endocrinol. 2013. PMID: 23211524 Free PMC article.
-
Peripubertal GnRH and testosterone co-treatment leads to increased familiarity preferences in male sheep.Psychoneuroendocrinology. 2019 Oct;108:70-77. doi: 10.1016/j.psyneuen.2019.06.008. Epub 2019 Jun 13. Psychoneuroendocrinology. 2019. PMID: 31229635 Free PMC article.
-
G protein-coupled receptors involved in GnRH regulation: molecular insights from human disease.Mol Cell Endocrinol. 2011 Oct 22;346(1-2):91-101. doi: 10.1016/j.mce.2011.06.022. Epub 2011 Jun 29. Mol Cell Endocrinol. 2011. PMID: 21736917 Free PMC article. Review.
-
Regulation of reproduction via tight control of gonadotropin hormone levels.Mol Cell Endocrinol. 2018 Mar 5;463:116-130. doi: 10.1016/j.mce.2017.03.022. Epub 2017 Mar 22. Mol Cell Endocrinol. 2018. PMID: 28342855 Free PMC article. Review.
-
GnRH regulation of Jun and Atf3 requires calcium, calcineurin, and NFAT.Mol Endocrinol. 2012 May;26(5):873-86. doi: 10.1210/me.2012-1045. Epub 2012 Mar 22. Mol Endocrinol. 2012. PMID: 22446101 Free PMC article.
References
-
- Armstrong S.P., Caunt C.J., Finch A.R., McArdle C.A. GnRH receptor signaling to ERK and NFAT: live cell imaging with sustained and pulsatile stimulation. 91st American Endocrine Society Meeting; Washington, DC, 1-6-2009; 2009.
-
- Bedecarrats G.Y., Kaiser U.B. Differential regulation of gonadotropin subunit gene promoter activity by pulsatile gonadotropin-releasing hormone (GnRH) in perifused L{beta}T2 cells: role of GnRH receptor concentration. Endocrinology. 2003;144:1802–1811. - PubMed
-
- Belchetz P.E., Plant T.M., Nakai Y., Keogh E.J., Knobil E. Hypophysial responses to continuous and intermittent delivery of hypopthalamic gonadotropin-releasing hormone. Science. 1978;202:631–633. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
