

Review
doi: 10.14310/horm.2002.1270.
Physiological effects and mechanisms of action of endocrine disrupting chemicals that alter estrogen signaling
Affiliations
- PMID: 20688617
- PMCID: PMC4782146
- DOI: 10.14310/horm.2002.1270
Item in Clipboard
Review
Physiological effects and mechanisms of action of endocrine disrupting chemicals that alter estrogen signaling
Hormones (Athens).
2010 Jul-Sep.
No abstract available
Figures
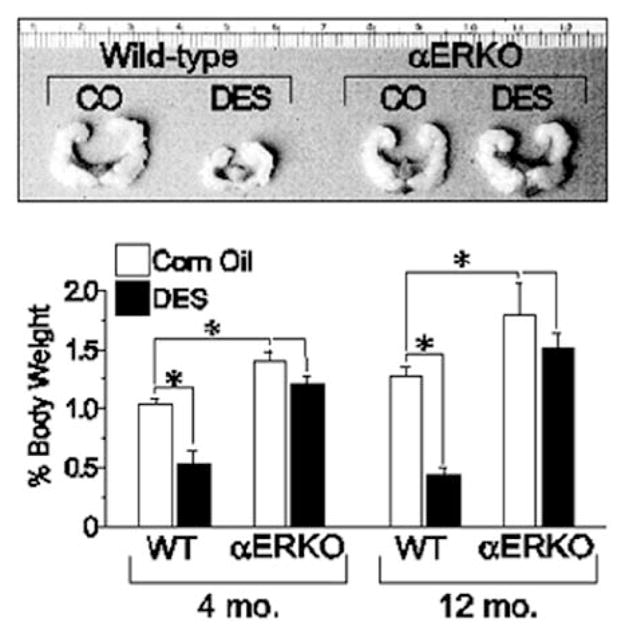
Effect of neonatal DES exposure on seminal vesicle weight in adult WT and αERKO mice. Top, seminal vesicles from 6-month old WT and αERKO mice following neonatal exposure to either vehicle (corn oil, CO) or DES (scale is in centimeters). Bottom, quantitative analysis of seminal vesicle weights from WT and aERKO mice at 4 and 12 months of age following neonatal exposure to CO or DES. *, P<0.05. (Modified from Prins, G.S., Birch, L., Couse, J.F., Choi, I., Katzenellenbogen, B., and Korach, K.S., 2001 Cancer Res., 61: 6089-6097.)
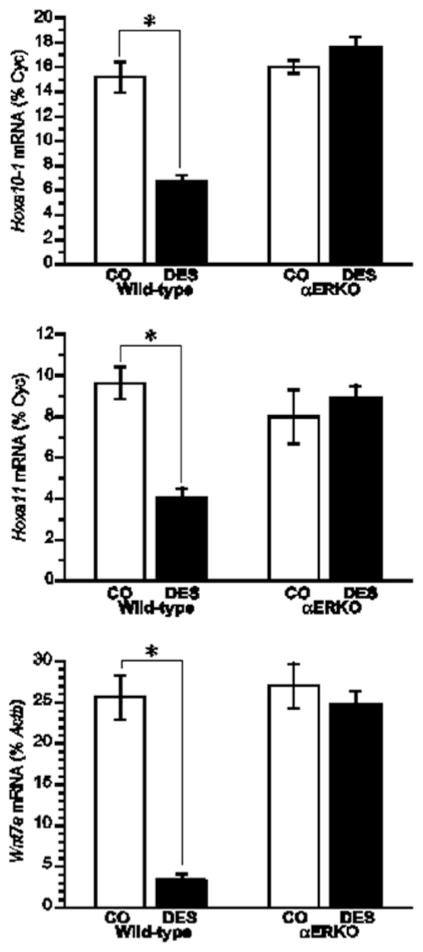
Effect of neonatal DES exposure on uterine expression of Hoxa10, Hoxa11, and Wnt7a in WT and αERKO mice. Top and middle, Quantitative analyses of ribonuclease protection assays (Hoxa10 and Hoxa11) (+/− SEM) showing average percentage of cyclophilin (normalization mRNA) for each treatment group. Bottom, Quantitative analysis of semiquantitative RT-PCR showing the average Wnt7a levels as a percentage of Actb (+/− SEM) for each treatment group. *, P<0.01. (Modified from Couse, J.F., Dixon, D., Yates, M., Moore, A.B., Ma, L., Maas, R., and Korach, K.S., 2001 Dev. Biol., 238: 224-238.).
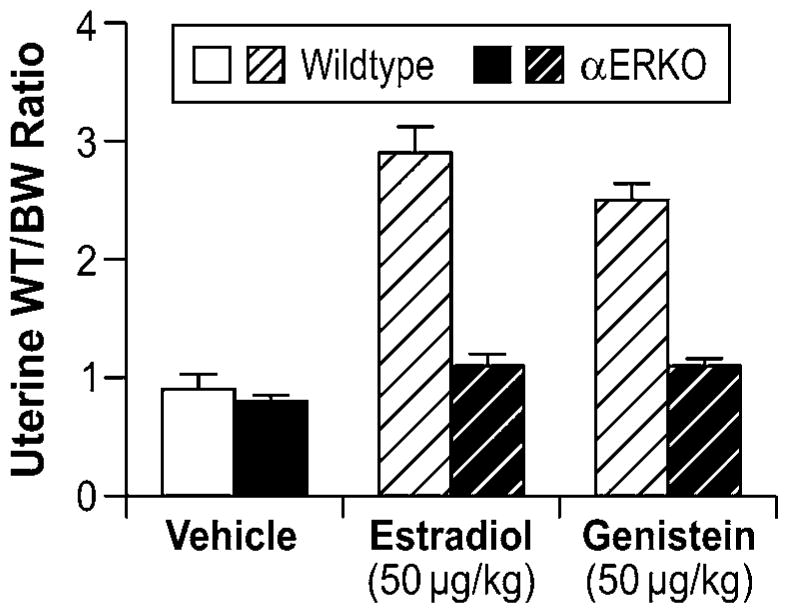
Role of ERα in modulating genistein-induced uterotropic effects. Ovariectomized WT and αERKO mice (n=4 mice/treatment group) were treated daily with subcutaneous injections of corn oil (vehicle), 10μg/kg estradiol, or 50μg/kg genistein for three days. On the fourth day whole body weights were measured and uteri were collected and weighed. The data represent uterine wet weight with respect to whole body weight. (Unpublished data from Lindzey, J. and Korach, K.S.).
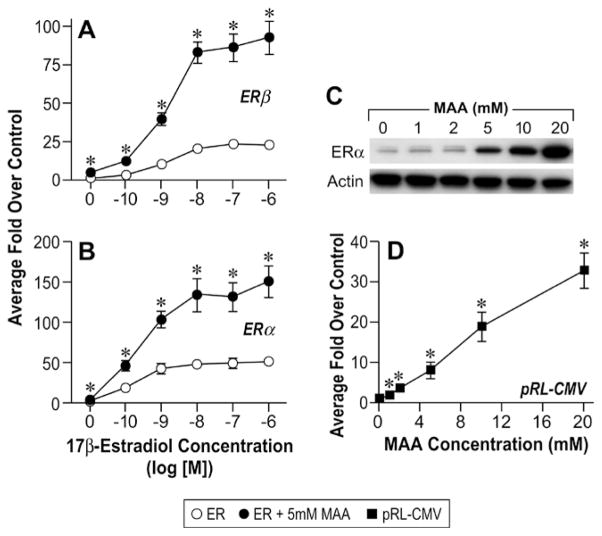
MAA activates the CMV promoter in vitro. A. HeLa cells were transiently transfected with a human ERβ expression vector, the estrogen-responsive 3X-ERE-TATA-Luc reporter plasmid, and the CMV-β-gal reporter plasmid and treated for 18h with either vehicle, increasing concentrations of E2, 5mM MAA, or increasing concentrations of E2 plus 5mM MAA. The histogram represents the average fold over control of duplicate samples from three independent experiments. The error bars represent the standard error of the mean (SEM), *p<0.05 comparing identical treatments +/− 5mM MAA. B. HeLa cells were transiently transfected with a human ERα expression vector, 3X-ERE-TATA-Luc and CMV-β-gal and treated identically to the cells in Figure 1A. The data shown represent the average fold over control of duplicate samples from three independent experiments and the error bars indicated the SEM, *p<0.05 comparing identical treatments +/− 5mM MAA. C. HeLa cells were transiently transfected with a human ERα expression vector and treated with either vehicle or increasing concentrations of MAA for 18h. Total protein was isolated from the cells and ERα protein expression was analyzed by Western blot. The data shown are representative of three independent experiments. D. HeLa cells were transiently transfected with the pRL-CMV reporter plasmid and treated with either vehicle or increasing concentrations of MAA for 18h. The data shown represent the average fold over control of duplicate samples from three independent experiments. The error bars represent the SEM, *p<0.01 comparing treatment with MAA to vehicle control. (Modified from Henley, D.V., Mueller, S., and Korach, K.S. In Press Environ Health Persp.).
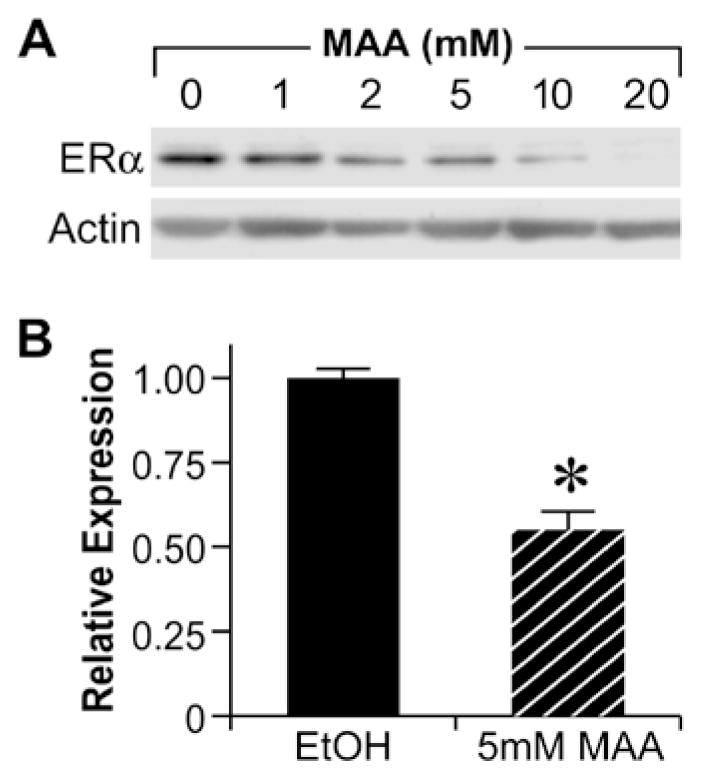
Methoxyacetic Acid (MAA) decreases endogenous ERα expression.. A. MCF-7 cells were treated for 24h with either vehicle or increasing concentrations of MAA. Total protein was isolated from each sample and analyzed for ERα protein expression by Western blot. The data shown are representative of results from three independent experiments. B. MCF-7 cells were treated for 24h with either vehicle or 5mM MAA. Total RNA was isolated from each sample and steady state mRNA levels of ERα were measured by real-time PCR. The data shown represent the average fold over control obtained from duplicate samples in four independent experiments, and the error bars indicate the SEM, *p<0.01 comparing treatment with 5mM MAA to vehicle control. (Modified from Henley, D.V., Mueller, S., and Korach, K.S. In Press Environ Health Persp.).
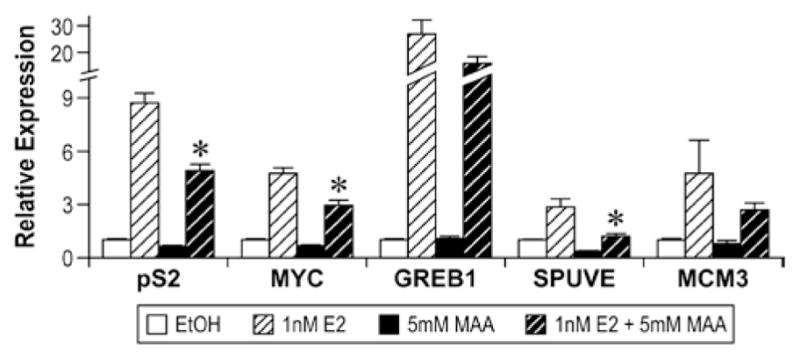
Methoxyacetic Acid (MAA) disrupts estrogen-mediated endogenous gene expression. MCF-7 cells were pretreated with either vehicle or 5mM MAA for 2h and then treated for 18h with either vehicle or 1nM E2. Total RNA was isolated from the samples and the expression of endogenous estrogen-responsive genes was analyzed by real-time PCR. The data shown represent the average fold over control of duplicate samples from at least three independent experiments. Error bars represent the SEM, *p<0.01 comparing treatment with 1nM E2 + 5mM MAA to treatment with 1nM E2 alone. The error bars represent SEM. (Modified from Henley, D.V., Mueller, S., and Korach, K.S. In Press Environ Health Persp.).
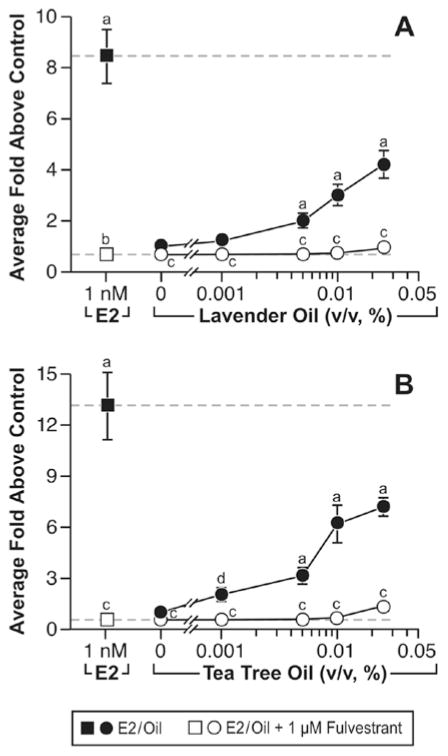
In vitro estrogenic activity of lavender oil and tea tree oil in reporter gene assays. MCF-7 cells were transiently transfected with the estrogen-inducible 3x-ERE-luciferase reporter plasmid and treated with increasing concentrations of either lavender oil or tea tree oil for 18 hours in the presence and absence of fulvestrant, an ER antagonist. Estradiol treatment (1 nM) was included as a positive control. Data are plotted as fold-luciferase activity over vehicle control. (Modified from Henley, D.V., Lipson, N., Korach, K.S., and Bloch, C.A., NEJM, 356, 481, 2007).
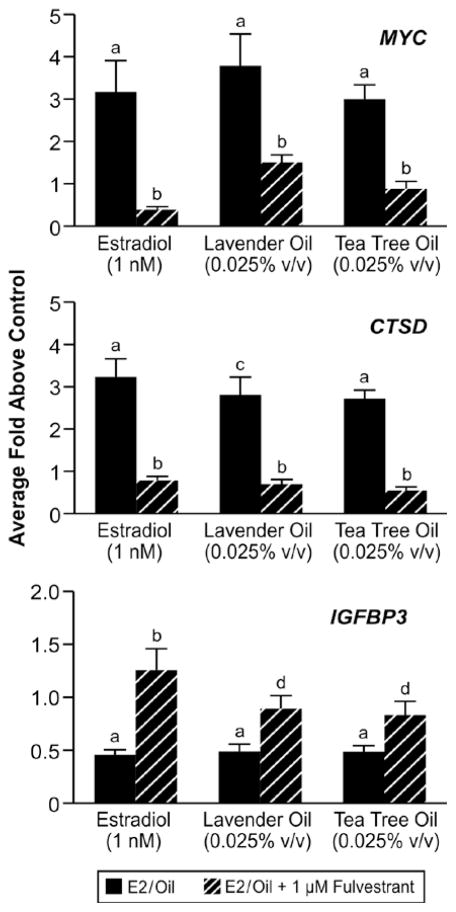
In vitro effects of lavender oil and tea tree oil on the expression of estrogen-regulated endogenous genes. MCF-7 cells were treated with 1 nM estradiol, 0.025% lavender oil, or 0.025% tea tree oil in the presence or absence of fulvestrant and real-time PCR was performed to measure changes in the expression of the estrogen-inducible genes MYC, IGFBP3, and CTSD. Data shown are normalized and relative to a vehicle control. (Modified from Henley, D.V., Lipson, N., Korach, K.S., and Bloch, C.A., NEJM, 356, 481, 2007).
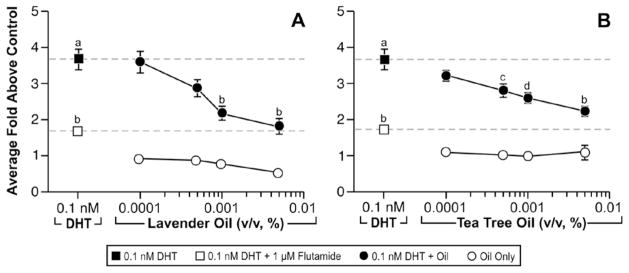
In vitro antiandrogenic activity of lavender oil and tea tree oil in reporter gene assays. MDA-kb2 cells stably transfected with the MMTV-luciferase reporter plasmid were treated for 24 hours with increasing concentrations of lavender oil or tea tree oil in the presence or absence of 0.1 nM dihydrotestosterone (DHT). The androgen receptor antagonist flutamide (1 μM) was included as a control antagonist for the androgen receptor. Luciferase activity was measured and data plotted as the average fold above control. (Modified from Henley, D.V., Lipson, N., Korach, K.S., and Bloch, C.A., NEJM, 356, 483, 2007).
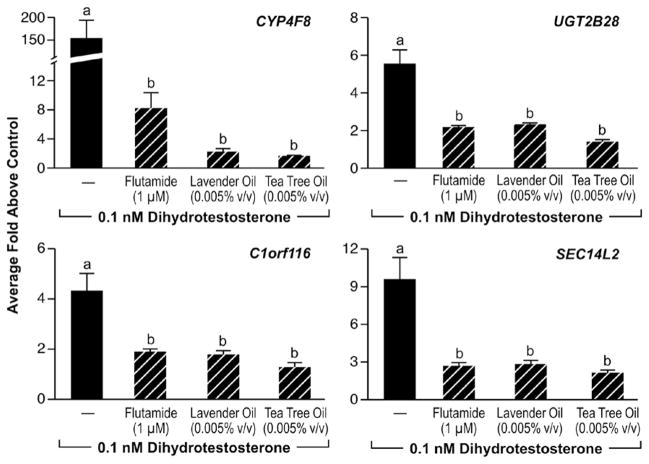
In vitro effects of lavender oil and tea tree oil on androgen-stimulated endogenous gene expression. MDA-kb2 cells were treated with 0.1 nM dihydrotestosterone for 24 hours in the presence or absence of 0.0005% lavender oil, 0.0005% tea tree oil, or 1μM flutamide and real-time PCR was performed to measure changes in the expression of androgen-inducible genes. Data shown are normalized and relative to a vehicle control. (Modified from Henley, D.V., Lipson, N., Korach, K.S., and Bloch, C.A., NEJM, 356, 483, 2007).
Similar articles
-
Endocrine-disrupting chemicals use distinct mechanisms of action to modulate endocrine system function.Endocrinology. 2006 Jun;147(6 Suppl):S25-32. doi: 10.1210/en.2005-1117. Epub 2006 May 11. Endocrinology. 2006. PMID: 16690802 Review.
-
Effects of Diethylstilbestrol on Zebrafish Gonad Development and Endocrine Disruption Mechanism.Biomolecules. 2021 Jun 25;11(7):941. doi: 10.3390/biom11070941. Biomolecules. 2021. PMID: 34201983 Free PMC article.
-
Chemicals with endocrine-disrupting potential: a threat to human health?Angew Chem Int Ed Engl. 2005 Sep 5;44(35):5568-74. doi: 10.1002/anie.200502138. Angew Chem Int Ed Engl. 2005. PMID: 16136609 No abstract available.
-
[Xenoestrogens: endocrine disrupting compounds].Ginekol Pol. 2008 Nov;79(11):785-90. Ginekol Pol. 2008. PMID: 19140503 Review. Polish.
-
Endocrine disrupting effects of zearalenone, alpha- and beta-zearalenol at the level of nuclear receptor binding and steroidogenesis.Toxicol Lett. 2011 Oct 10;206(2):210-7. doi: 10.1016/j.toxlet.2011.07.015. Epub 2011 Jul 23. Toxicol Lett. 2011. PMID: 21803136
Cited by
-
Differential in Vitro Biological Action, Coregulator Interactions, and Molecular Dynamic Analysis of Bisphenol A (BPA), BPAF, and BPS Ligand-ERα Complexes.Environ Health Perspect. 2018 Jan 31;126(1):017012. doi: 10.1289/EHP2505. Environ Health Perspect. 2018. PMID: 29389661 Free PMC article.
-
Drugs and Medical Devices: Adverse Events and the Impact on Women's Health.Clin Ther. 2017 Jan;39(1):10-22. doi: 10.1016/j.clinthera.2016.12.009. Epub 2017 Jan 7. Clin Ther. 2017. PMID: 28069260 Free PMC article. Review.
-
Fingerprinting of neurotoxic compounds using a mouse embryonic stem cell dual luminescence reporter assay.Arch Toxicol. 2017 Jan;91(1):365-391. doi: 10.1007/s00204-016-1690-2. Epub 2016 Mar 25. Arch Toxicol. 2017. PMID: 27015953 Free PMC article.
-
Activation of Estrogen Receptor by Bavachin from Psoralea corylifolia.Biomol Ther (Seoul). 2012 Mar;20(2):183-8. doi: 10.4062/biomolther.2012.20.2.183. Biomol Ther (Seoul). 2012. PMID: 24116293 Free PMC article.
-
Estrogen receptor is activated by korean red ginseng in vitro but not in vivo.J Ginseng Res. 2012 Apr;36(2):169-75. doi: 10.5142/jgr.2012.36.2.169. J Ginseng Res. 2012. PMID: 23717117 Free PMC article.
References
-
- Dodds EC, Lawsaon W, Robinson R. Estrogenic activity of certain synthetic compounds. Nature. 1938:141247–141248.
-
- Smith OW, Smith GBS, Hurwitz D. Increased excretion of pregnanediol in pregnancy from diethylstilbestrol with special reference to the prevention of late pregnancy accidents. Med Rec Ann. 1946;40:1669–1671. - PubMed
-
- Dieckmann WJ, Davis ME, Rynkiewicz LM, Pottinger RE, Gabbe SG. Does the administration of diethylstilbestrol during pregnancy have therapeutic value 1953? Am J Obstet Gynecol. 1999;66:1062–1081. - PubMed
-
- Herbst AL, Ulfelder H, Poskanzer DC. Adenocarcinoma of the vagina. Association of maternal stilbestrol therapy with tumor appearance in young women. N Engl J Med. 1971;284:878–881. - PubMed
-
- Melnick S, Cole P, Anderson D, Herbst A. Rates and risks of diethylstilbestrol-related clear-cell adenocarcinoma of the vagina and cervix. An update. N Engl J Med. 1987;316:514–516. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
