

Uptake of selenite by Saccharomyces cerevisiae involves the high and low affinity orthophosphate transporters
- PMID: 20688911
- PMCID: PMC2952204
- DOI: 10.1074/jbc.M110.139865
Uptake of selenite by Saccharomyces cerevisiae involves the high and low affinity orthophosphate transporters
Abstract
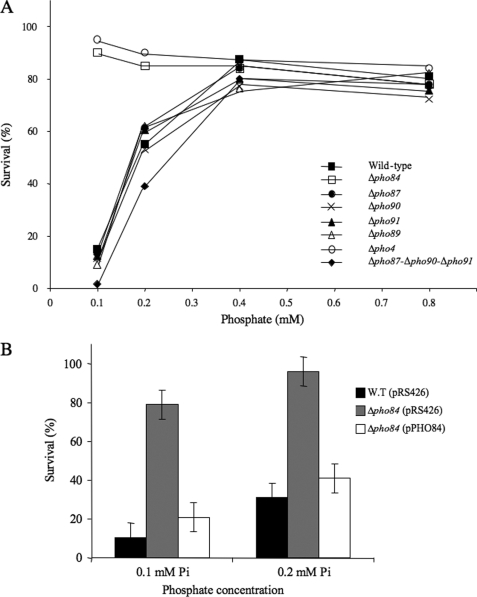
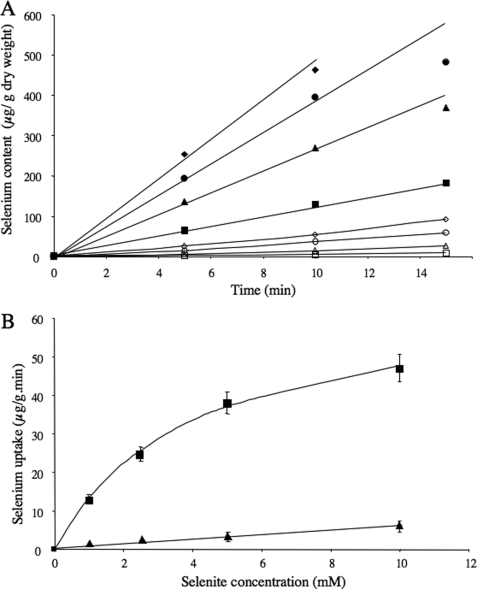
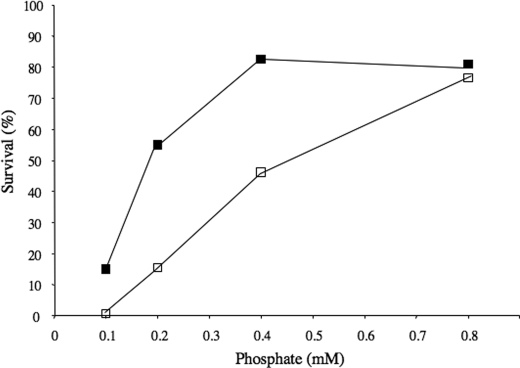
Although the general cytotoxicity of selenite is well established, the mechanism by which this compound crosses cellular membranes is still unknown. Here, we show that in Saccharomyces cerevisiae, the transport system used opportunistically by selenite depends on the phosphate concentration in the growth medium. Both the high and low affinity phosphate transporters are involved in selenite uptake. When cells are grown at low P(i) concentrations, the high affinity phosphate transporter Pho84p is the major contributor to selenite uptake. When phosphate is abundant, selenite is internalized through the low affinity P(i) transporters (Pho87p, Pho90p, and Pho91p). Accordingly, inactivation of the high affinity phosphate transporter Pho84p results in increased resistance to selenite and reduced uptake in low P(i) medium, whereas deletion of SPL2, a negative regulator of low affinity phosphate uptake, results in exacerbated sensitivity to selenite. Measurements of the kinetic parameters for selenite and phosphate uptake demonstrate that there is a competition between phosphate and selenite ions for both P(i) transport systems. In addition, our results indicate that Pho84p is very selective for phosphate as compared with selenite, whereas the low affinity transporters discriminate less efficiently between the two ions. The properties of phosphate and selenite transport enable us to propose an explanation to the paradoxical increase of selenite toxicity when phosphate concentration in the growth medium is raised above 1 mm.
Figures


Similar articles
-
The Saccharomyces cerevisiae high affinity phosphate transporter encoded by PHO84 also functions in manganese homeostasis.J Biol Chem. 2003 Oct 24;278(43):42036-40. doi: 10.1074/jbc.M307413200. Epub 2003 Aug 15. J Biol Chem. 2003. PMID: 12923174
-
The competitive advantage of a dual-transporter system.Science. 2011 Dec 9;334(6061):1408-12. doi: 10.1126/science.1207154. Science. 2011. PMID: 22158820 Free PMC article.
-
The yeast Aft2 transcription factor determines selenite toxicity by controlling the low affinity phosphate transport system.Sci Rep. 2016 Sep 13;6:32836. doi: 10.1038/srep32836. Sci Rep. 2016. PMID: 27618952 Free PMC article.
-
The conservation of phosphate-binding residues among PHT1 transporters suggests that distinct transport affinities are unlikely to result from differences in the phosphate-binding site.Biochem Soc Trans. 2016 Oct 15;44(5):1541-1548. doi: 10.1042/BST20160016. Biochem Soc Trans. 2016. PMID: 27911737 Review.
-
Inorganic Phosphate and Sulfate Transport in S. cerevisiae.Adv Exp Med Biol. 2016;892:253-269. doi: 10.1007/978-3-319-25304-6_10. Adv Exp Med Biol. 2016. PMID: 26721277 Review.
Cited by
-
Selenium cycling across soil-plant-atmosphere interfaces: a critical review.Nutrients. 2015 May 29;7(6):4199-239. doi: 10.3390/nu7064199. Nutrients. 2015. PMID: 26035246 Free PMC article. Review.
-
Metabolic Response of the Yeast Candida utilis During Enrichment in Selenium.Int J Mol Sci. 2020 Jul 25;21(15):5287. doi: 10.3390/ijms21155287. Int J Mol Sci. 2020. PMID: 32722488 Free PMC article.
-
Equilibrium modeling of selenium binding from aqueous solutions by Candida utilis ATCC 9950 yeasts.3 Biotech. 2018 Sep;8(9):388. doi: 10.1007/s13205-018-1415-8. Epub 2018 Aug 27. 3 Biotech. 2018. PMID: 30175025 Free PMC article.
-
Comparative transcriptomics provides novel insights into the mechanisms of selenium accumulation and transportation in tea cultivars (Camellia sinensis (L.) O. Kuntze).Front Plant Sci. 2023 Oct 2;14:1268537. doi: 10.3389/fpls.2023.1268537. eCollection 2023. Front Plant Sci. 2023. PMID: 37849840 Free PMC article.
-
Kratosvirus quantuckense: the history and novelty of an algal bloom disrupting virus and a model for giant virus research.Front Microbiol. 2023 Nov 30;14:1284617. doi: 10.3389/fmicb.2023.1284617. eCollection 2023. Front Microbiol. 2023. PMID: 38098665 Free PMC article. Review.
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
