

A short receptor downregulates JAK/STAT signalling to control the Drosophila cellular immune response
- PMID: 20689801
- PMCID: PMC2914635
- DOI: 10.1371/journal.pbio.1000441
A short receptor downregulates JAK/STAT signalling to control the Drosophila cellular immune response
Abstract
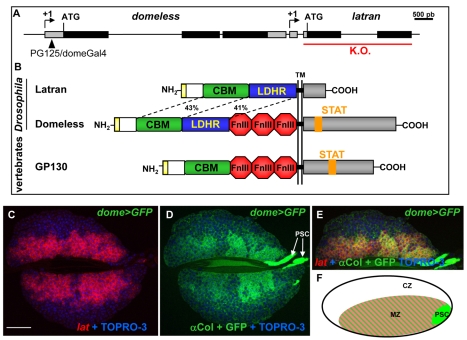
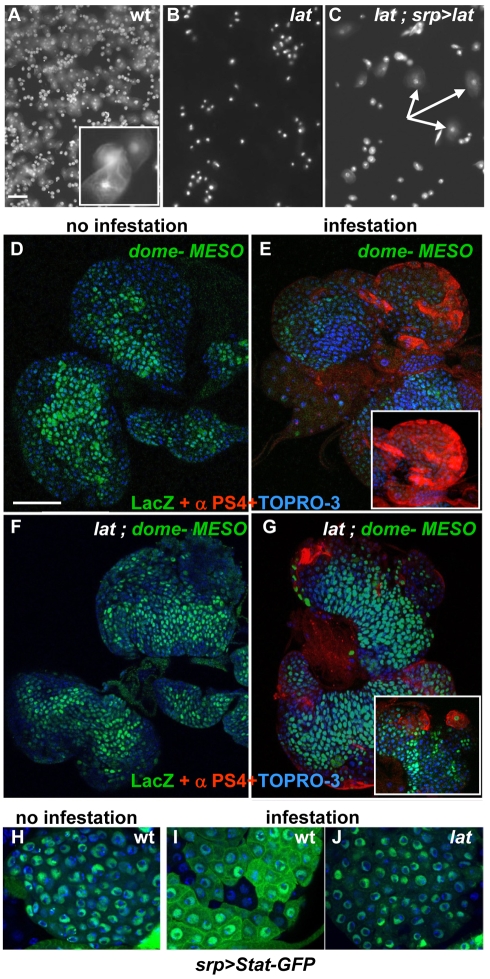
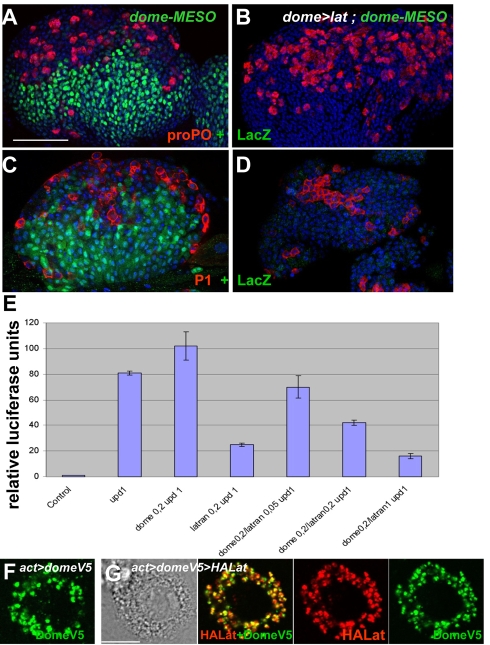
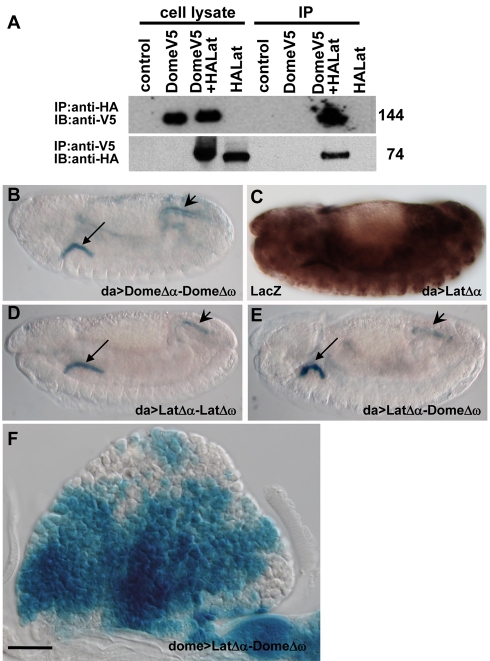
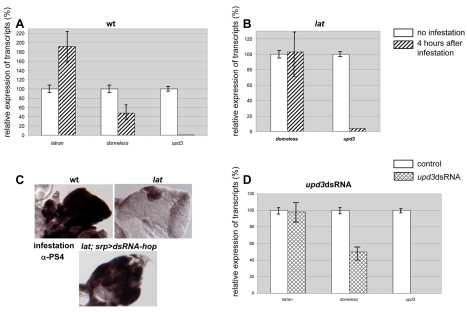
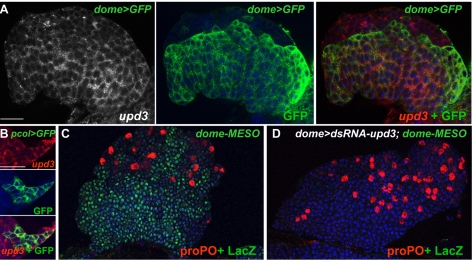
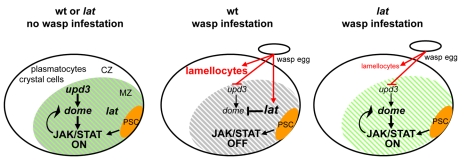
The posterior signalling centre (PSC), a small group of specialised cells, controls hemocyte (blood cell) homeostasis in the Drosophila larval hematopoietic organ, the lymph gland. This role of the PSC is very reminiscent of the "niche," the micro-environment of hematopoietic stem cells in vertebrates. We have recently shown that the PSC acts in a non-cell-autonomous manner to maintain janus tyrosine kinase/signal transducers and activators of transcription (JAK/STAT) signalling in hematopoietic progenitors (prohemocytes), thereby preserving the multipotent character necessary for their differentiation into lamellocytes, a cryptic and dedicated immune cell type required to fight specific immune threats such as wasp parasitism. In this report, on the basis of a knock out generated by homologous recombination, we show that a short type I cytokine-related receptor CG14225/Latran is required for switching off JAK/STAT signalling in prohemocytes. This is a prerequisite to massive differentiation of lamellocytes upon wasp parasitisation. In vivo and cell culture assays indicate that Latran forms heteromers with Domeless, the Drosophila type I cytokine signalling receptor related to mammalian GP130, and antagonises Domeless activity in a dose-dependent manner. Our analysis further shows that a primary immune response to wasp parasitism is a strong decrease in cytokine mRNA levels in the lymph gland, followed by an increase in the latran/domeless ratio. We propose that this sequence of events culminates in the complete inhibition of residual JAK/STAT signalling by Latran. JAK/STAT activity has been associated with several human diseases including leukaemia while knock-out studies in mice point to a central role of this pathway in hematopoiesis and regulation of immune functions. The specific function of Drosophila Latran is, to our knowledge, the first in vivo example of a role for a nonsignalling receptor in controlling a dedicated immune response, and thus raises the question of whether short, nonsignalling receptors also control specific aspects of vertebrate cellular immunity.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Control of blood cell homeostasis in Drosophila larvae by the posterior signalling centre.Nature. 2007 Mar 15;446(7133):325-8. doi: 10.1038/nature05650. Nature. 2007. PMID: 17361184
-
Differential activation of JAK-STAT signaling reveals functional compartmentalization in Drosophila blood progenitors.Elife. 2021 Feb 17;10:e61409. doi: 10.7554/eLife.61409. Elife. 2021. PMID: 33594977 Free PMC article.
-
Mechanical stress to Drosophila larvae stimulates a cellular immune response through the JAK/STAT signaling pathway.Biochem Biophys Res Commun. 2018 Jul 20;502(3):415-421. doi: 10.1016/j.bbrc.2018.05.192. Epub 2018 May 30. Biochem Biophys Res Commun. 2018. PMID: 29856996
-
What a tangled web we weave: crosstalk between JAK-STAT and other signalling pathways during development in Drosophila.FEBS J. 2025 Jul;292(13):3298-3320. doi: 10.1111/febs.17391. Epub 2025 Jan 16. FEBS J. 2025. PMID: 39821459 Review.
-
Ontogeny of the Drosophila larval hematopoietic organ, hemocyte homeostasis and the dedicated cellular immune response to parasitism.Int J Dev Biol. 2010;54(6-7):1117-25. doi: 10.1387/ijdb.093053jk. Int J Dev Biol. 2010. PMID: 20711989 Review.
Cited by
-
Drosophila as a Genetic Model for Hematopoiesis.Genetics. 2019 Feb;211(2):367-417. doi: 10.1534/genetics.118.300223. Genetics. 2019. PMID: 30733377 Free PMC article. Review.
-
Two Independent Functions of Collier/Early B Cell Factor in the Control of Drosophila Blood Cell Homeostasis.PLoS One. 2016 Feb 11;11(2):e0148978. doi: 10.1371/journal.pone.0148978. eCollection 2016. PLoS One. 2016. PMID: 26866694 Free PMC article.
-
Obligate symbionts activate immune system development in the tsetse fly.J Immunol. 2012 Apr 1;188(7):3395-403. doi: 10.4049/jimmunol.1103691. Epub 2012 Feb 24. J Immunol. 2012. PMID: 22368278 Free PMC article.
-
Regulation of phagocyte triglyceride by a STAT-ATG2 pathway controls mycobacterial infection.Nat Commun. 2017 Mar 6;8:14642. doi: 10.1038/ncomms14642. Nat Commun. 2017. PMID: 28262681 Free PMC article.
-
Serotonergic neurons regulate the Drosophila vascular niche to control immune stress hematopoiesis.Nat Commun. 2025 Jun 3;16(1):5152. doi: 10.1038/s41467-025-60493-y. Nat Commun. 2025. PMID: 40461546 Free PMC article.
References
-
- Lemaitre B, Hoffmann J. The host defense of Drosophila melanogaster. Annu Rev Immunol. 2007;25:697–743. - PubMed
-
- Crozatier M, Meister M. Drosophila haematopoiesis. Cell Microbiol. 2007;9:1117–1126. - PubMed
-
- Evans C. J, Banerjee U. Transcriptional regulation of hematopoiesis in Drosophila. Blood Cells Mol Dis. 2003;30:223–228. - PubMed
-
- Bataille L, Auge B, Ferjoux G, Haenlin M, Waltzer L. Resolving embryonic blood cell fate choice in Drosophila: interplay of GCM and RUNX factors. Development. 2005;132:4635–4644. - PubMed
-
- Lanot R, Zachary D, Holder F, Meister M. Postembryonic hematopoiesis in Drosophila. Dev Biol. 2001;230:243–257. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
