

Sequential activation of apical and basolateral contractility drives ascidian endoderm invagination
- PMID: 20691592
- PMCID: PMC4088275
- DOI: 10.1016/j.cub.2010.06.075
Sequential activation of apical and basolateral contractility drives ascidian endoderm invagination
Abstract
Background: Epithelial invagination is a fundamental morphogenetic behavior that transforms a flat cell sheet into a pit or groove. Previous studies of invagination have focused on the role of actomyosin-dependent apical contraction; other mechanisms remain largely unexplored.
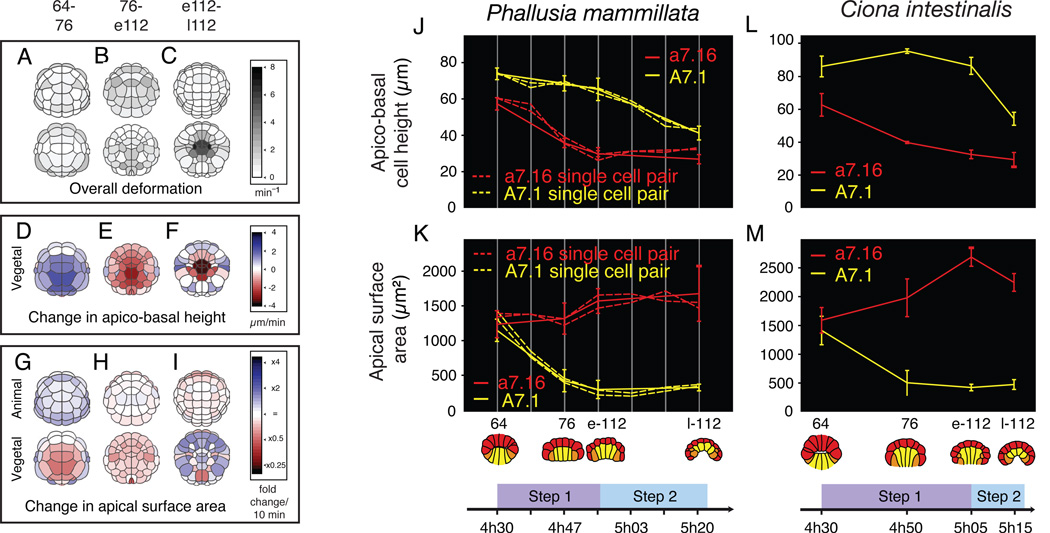
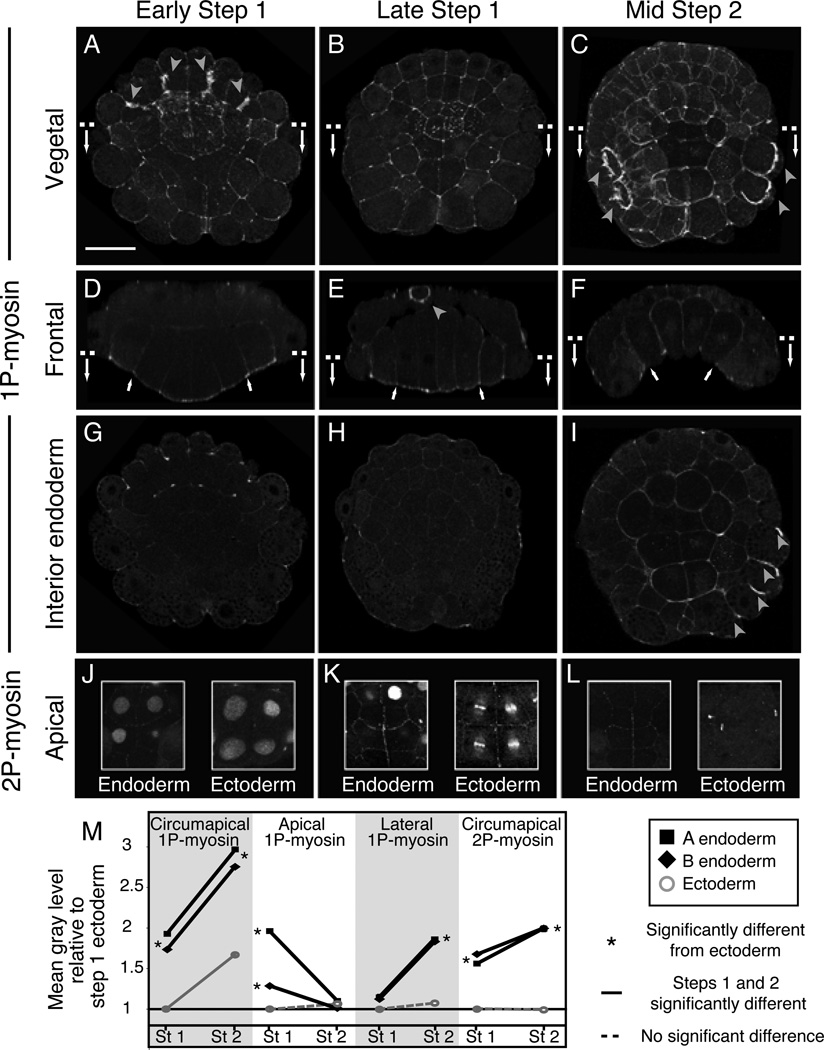
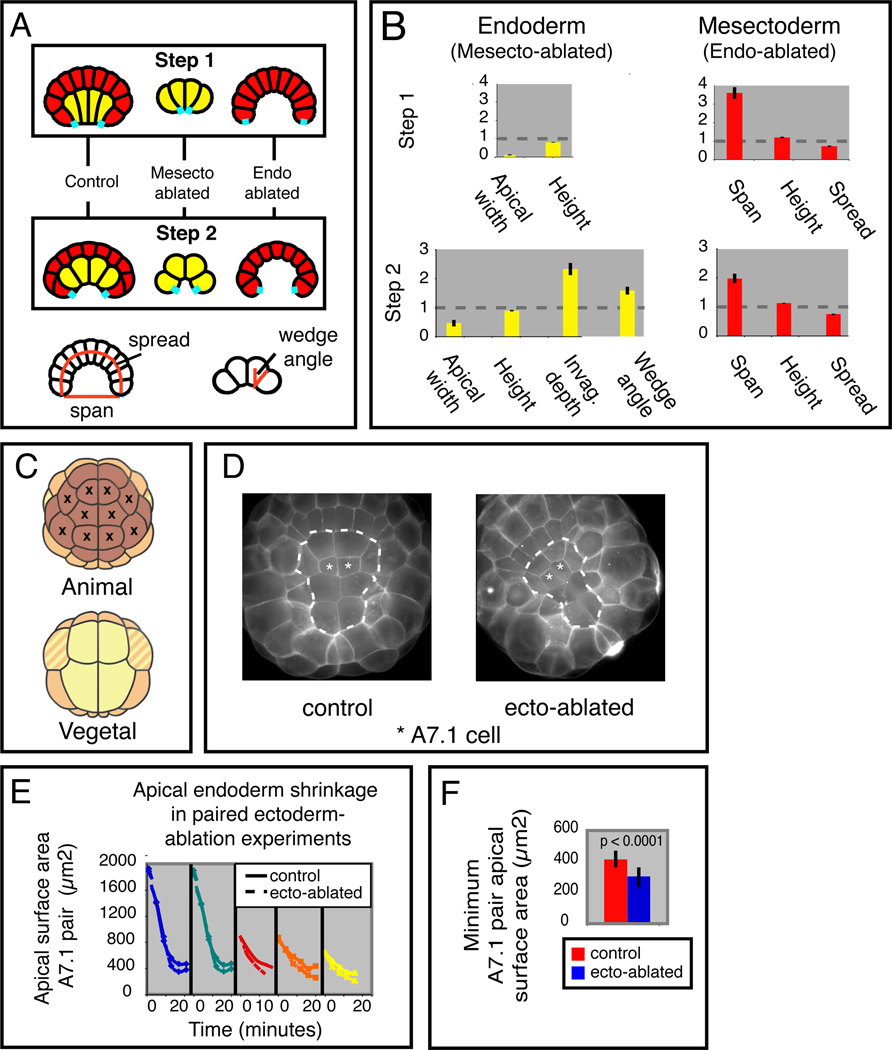
Results: We combined experimental and computational approaches to identify a two-step mechanism for endoderm invagination during ascidian gastrulation. During Step 1, which immediately precedes invagination, endoderm cells constrict their apices because of Rho/Rho-kinase-dependent apical enrichment of 1P-myosin. Our data suggest that endoderm invagination itself occurs during Step 2, without further apical shrinkage, via a novel mechanism we call collared rounding: Rho/Rho-kinase-independent basolateral enrichment of 1P-myosin drives apico-basal shortening, whereas Rho/Rho-kinase-dependent enrichment of 1P and 2P myosin in circumapical collars is required to prevent apical expansion and for deep invagination. Simulations show that boundary-specific tension values consistent with these distributions of active myosin can explain the cell shape changes observed during invagination both in normal embryos and in embryos treated with pharmacological inhibitors of either Rho-kinase or Myosin II ATPase. Indeed, we find that the balance of strong circumapical and basolateral tension is the only mechanism based on differential cortical tension that can explain ascidian endoderm invagination. Finally, simulations suggest that mesectoderm cells resist endoderm shape changes during both steps, and we confirm this prediction experimentally.
Conclusions: Our findings suggest that early ascidian gastrulation is driven by the coordinated apposition of circumapical and lateral endoderm contraction, working against a resisting mesectoderm. We propose that similar mechanisms may operate during other invaginations.
Copyright © 2010 Elsevier Ltd. All rights reserved.
Figures




References
-
- Davidson L, Koehl MAR, Keller R, Oster GF. How do sea urchins invaginate? Using biomechanics to distinguish between mechanisms of primary invagination. Development. 1995;121:2005–2018. - PubMed
-
- Keller , Davidson RL, Shook D. How we are shaped: The biomechanics of gastrulation. Differentiation. 2003;71:171–205. - PubMed
-
- Clausi DA, Brodland GW. Mechanical evaluation of theories of neurulation using computer simulations. Development. 1993;118:1013–1023.
-
- Jacobsen AG, Oster GF, Odell GM, Cheng LY. Neurulation and the cortical tractor model for epithelial folding. Journal of embryology and experimental morphology. 1986;96:19–49. - PubMed
-
- Muñoz JJ, Barrett K, Miodownik M. A deformation gradient decomposition method for the analysis of the mechanics of morphogenesis. Journal of Biomechanics. 2007;40:1372–1380. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
