

Bicaudal-D regulates fragile X mental retardation protein levels, motility, and function during neuronal morphogenesis
- PMID: 20691595
- PMCID: PMC2927779
- DOI: 10.1016/j.cub.2010.07.016
Bicaudal-D regulates fragile X mental retardation protein levels, motility, and function during neuronal morphogenesis
Abstract
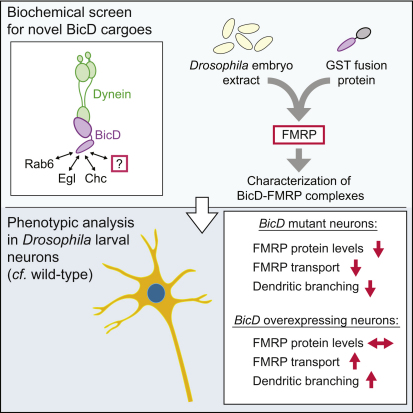
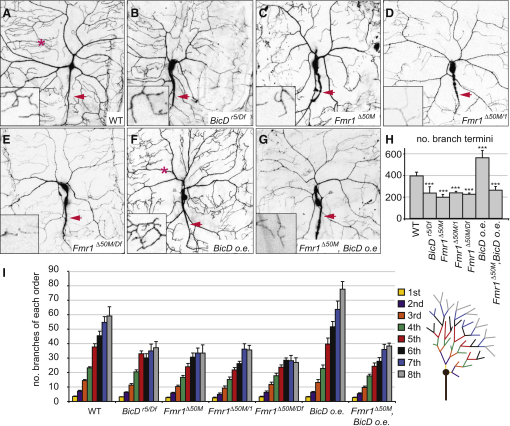
The expression of the RNA-binding factor Fragile X mental retardation protein (FMRP) is disrupted in the most common inherited form of cognitive deficiency in humans. FMRP controls neuronal morphogenesis by mediating the translational regulation and localization of a large number of mRNA targets, and these functions are closely associated with transport of FMRP complexes within neurites by microtubule-based motors. However, the mechanisms that link FMRP to motors and regulate its transport are poorly understood. Here we show that FMRP is complexed with Bicaudal-D (BicD) through a domain in the latter protein that mediates linkage of cargoes with the minus-end-directed motor dynein. We demonstrate in Drosophila that the motility and, surprisingly, levels of FMRP protein are dramatically reduced in BicD mutant neurons, leading to a paucity of FMRP within processes. We also provide functional evidence that BicD and FMRP cooperate to control dendritic morphogenesis in the larval nervous system. Our findings open new perspectives for understanding localized mRNA functions in neurons.
Copyright 2010 Elsevier Ltd. All rights reserved.
Figures



References
-
- Estes P.S., O'Shea M., Clasen S., Zarnescu D.C. Fragile X protein controls the efficacy of mRNA transport in Drosophila neurons. Mol. Cell. Neurosci. 2008;39:170–179. - PubMed
-
- Davidovic L., Jaglin X.H., Lepagnol-Bestel A.M., Tremblay S., Simonneau M., Bardoni B., Khandjian E.W. The fragile X mental retardation protein is a molecular adaptor between the neurospecific KIF3C kinesin and dendritic RNA granules. Hum. Mol. Genet. 2007;16:3047–3058. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
