

Eisosome organization in the filamentous ascomycete Aspergillus nidulans
- PMID: 20693301
- PMCID: PMC2950425
- DOI: 10.1128/EC.00087-10
Eisosome organization in the filamentous ascomycete Aspergillus nidulans
Abstract
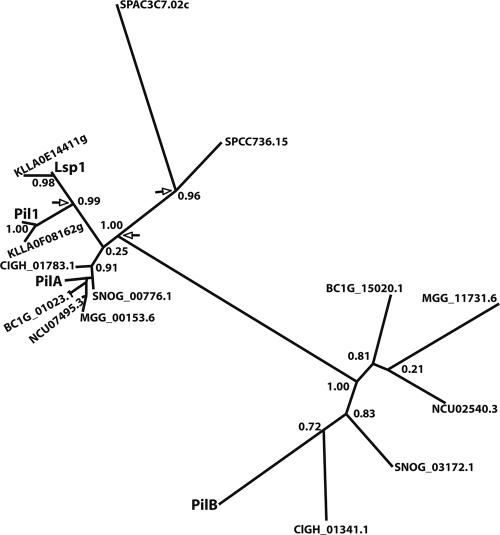
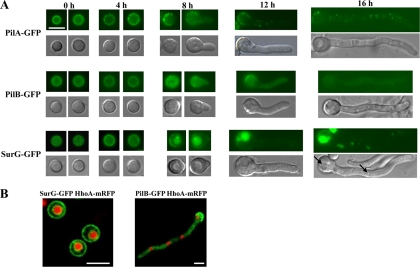
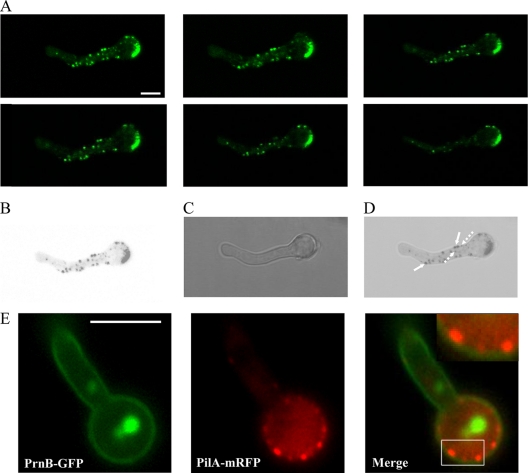
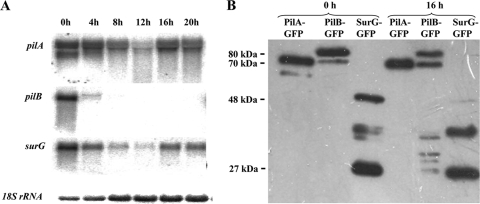
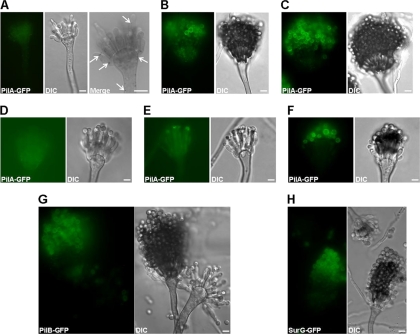
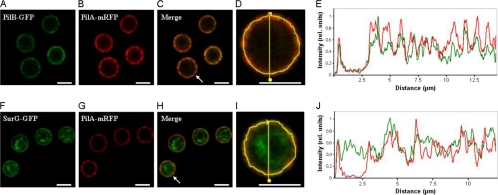
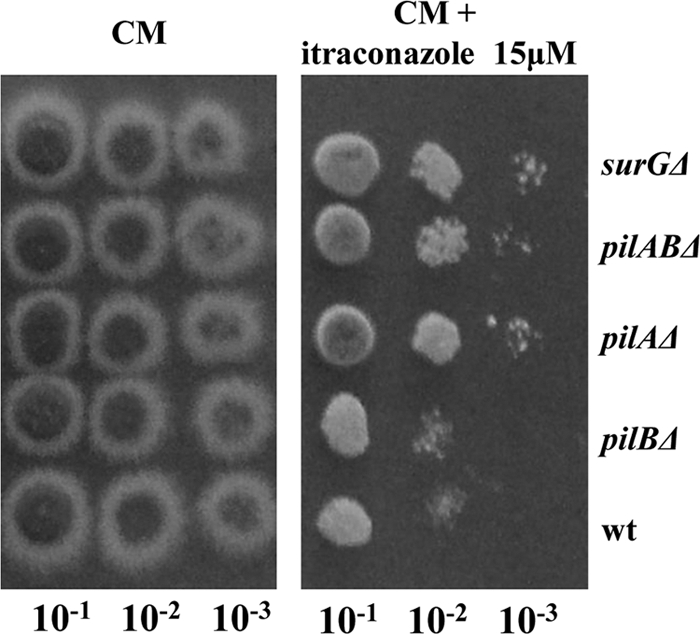
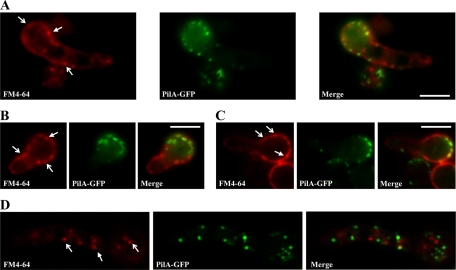
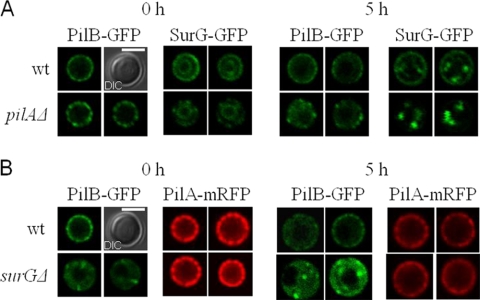
Eisosomes are subcortical organelles implicated in endocytosis and have hitherto been described only in Saccharomyces cerevisiae. They comprise two homologue proteins, Pil1 and Lsp1, which colocalize with the transmembrane protein Sur7. These proteins are universally conserved in the ascomycetes. We identify in Aspergillus nidulans (and in all members of the subphylum Pezizomycotina) two homologues of Pil1/Lsp1, PilA and PilB, originating from a duplication independent from that extant in the subphylum Saccharomycotina. In the aspergilli there are several Sur7-like proteins in each species, including one strict Sur7 orthologue (SurG in A. nidulans). In A. nidulans conidiospores, but not in hyphae, the three proteins colocalize at the cell cortex and form tightly packed punctate structures that appear different from the clearly distinct eisosome patches observed in S. cerevisiae. These structures are assembled late during the maturation of conidia. In mycelia, punctate structures are present, but they are composed only of PilA, while PilB is diffused in the cytoplasm and SurG is located in vacuoles and endosomes. Deletion of each of the genes does not lead to any obvious growth phenotype, except for moderate resistance to itraconazole. We could not find any obvious association between mycelial (PilA) eisosome-like structures and endocytosis. PilA and SurG are necessary for conidial eisosome organization in ways that differ from those for their S. cerevisiae homologues. These data illustrate that conservation of eisosomal proteins within the ascomycetes is accompanied by a striking functional divergence.
Figures
References
-
- Abenza J. F., Pantazopoulou A., Rodríguez J. M., Galindo A., Peñalva M. A. 2009. Long-distance movement of Aspergillus nidulans early endosomes on microtubule tracks. Traffic 10:57–75 - PubMed
-
- Anisimova M., Gascuel O. 2006. Approximate likelihood-ratio test for branches: a fast, accurate, and powerful alternative. Syst. Biol. 55:539–552 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
