

Setaria viridis: a model for C4 photosynthesis
- PMID: 20693355
- PMCID: PMC2947182
- DOI: 10.1105/tpc.110.075309
Setaria viridis: a model for C4 photosynthesis
Abstract
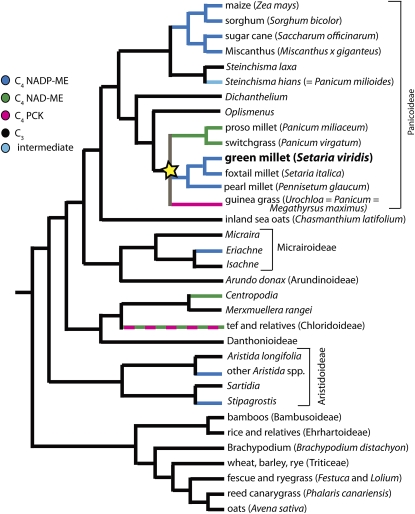
C(4) photosynthesis drives productivity in several major food crops and bioenergy grasses, including maize (Zea mays), sugarcane (Saccharum officinarum), sorghum (Sorghum bicolor), Miscanthus x giganteus, and switchgrass (Panicum virgatum). Gains in productivity associated with C(4) photosynthesis include improved water and nitrogen use efficiencies. Thus, engineering C(4) traits into C(3) crops is an attractive target for crop improvement. However, the lack of a small, rapid cycling genetic model system to study C(4) photosynthesis has limited progress in dissecting the regulatory networks underlying the C(4) syndrome. Setaria viridis is a member of the Panicoideae clade and is a close relative of several major feed, fuel, and bioenergy grasses. It is a true diploid with a relatively small genome of ~510 Mb. Its short stature, simple growth requirements, and rapid life cycle will greatly facilitate genetic studies of the C(4) grasses. Importantly, S. viridis uses an NADP-malic enzyme subtype C(4) photosynthetic system to fix carbon and therefore is a potentially powerful model system for dissecting C(4) photosynthesis. Here, we summarize some of the recent advances that promise greatly to accelerate the use of S. viridis as a genetic system. These include our recent successful efforts at regenerating plants from seed callus, establishing a transient transformation system, and developing stable transformation.
Figures

References
-
- Brown N.J., Parsley K., Hibberd J.M. (2005). The future of C4 research–Maize, Flaveria or Cleome? Trends Plant Sci. 10: 215–221 - PubMed
-
- Christin P.A., Besnard G., Samaritani E., Duvall M.R., Hodkinson T.R., Savolainen V., Salamin N. (2008). Oligocene CO2 decline promoted C4 photosynthesis in grasses. Curr. Biol. 18: 37–43 - PubMed
-
- Dekker J. (2003). Evolutionary biology of the foxtail (Setaria) species-group. Weed Biology and Management, Inderjit, ed (Dordrecht, The Netherlands: Kluwer; ), pp. 65–114
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
