

Toll-like receptor agonists: are they good adjuvants?
- PMID: 20693851
- PMCID: PMC2922045
- DOI: 10.1097/PPO.0b013e3181eaca65
Toll-like receptor agonists: are they good adjuvants?
Abstract
Therapeutic immunization leading to cancer regression remains a significant challenge. Successful immunization requires activation of adaptive immunity, including tumor specific CD4 T cells and CD8 T cells. Generally, the activation of T cells is compromised in patients with cancer because of immune suppression, loss of tumor antigen expression, and dysfunction of antigen-presenting cells. Antigen-presenting cells such as dendritic cells (DCs) are key for the induction of adaptive antitumor immune responses. Recently, attention has focused on novel adjuvants that enhance dendritic cell function and their ability to prime T cells. Agonists that target toll-like receptors are being used clinically either alone or in combination with tumor antigens and showing initial success both in terms of enhancing immune responses and eliciting antitumor activity. This review summarizes the application of these adjuvants to treat cancer and the potential for boosting responses in vivo.
Conflict of interest statement
Conflict of Interest Statement:
All authors have no conflict of interest to declare.
Figures
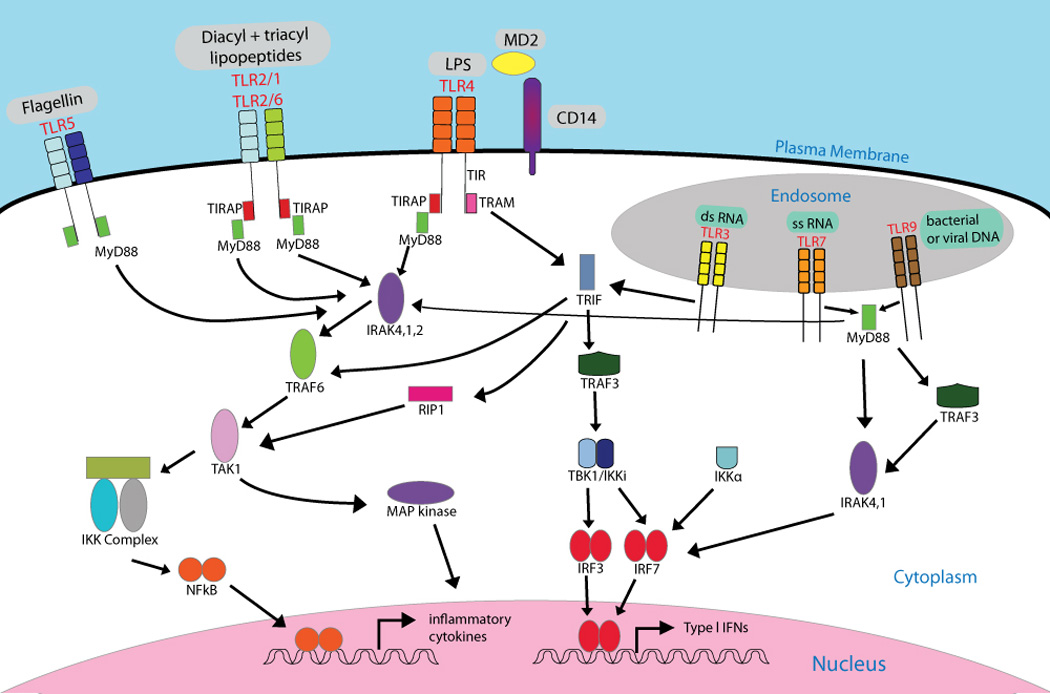
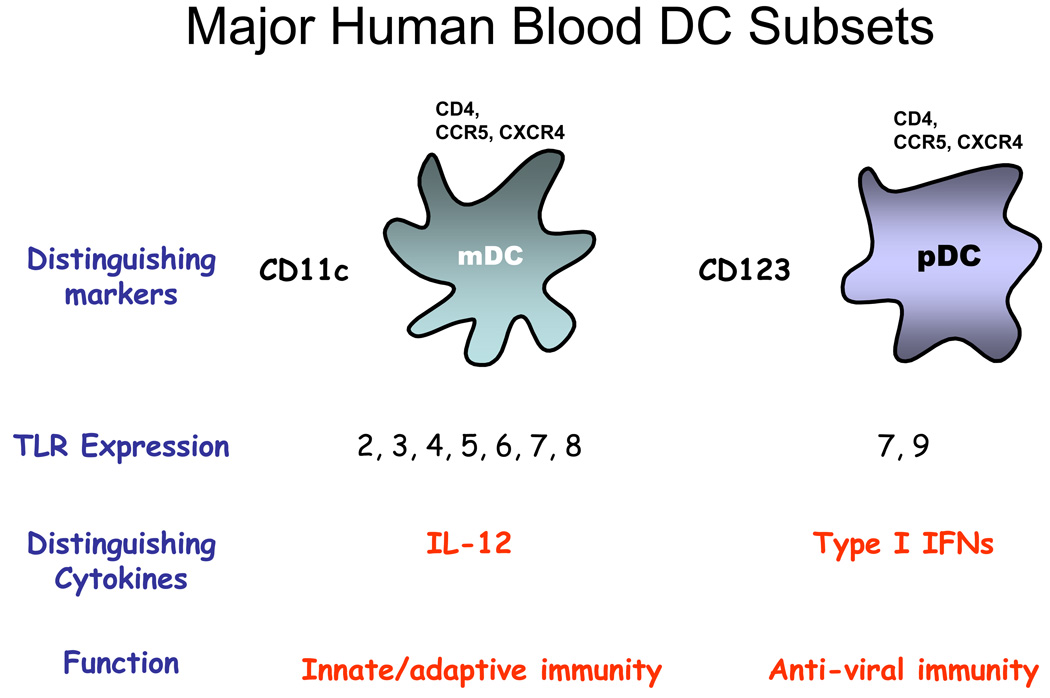
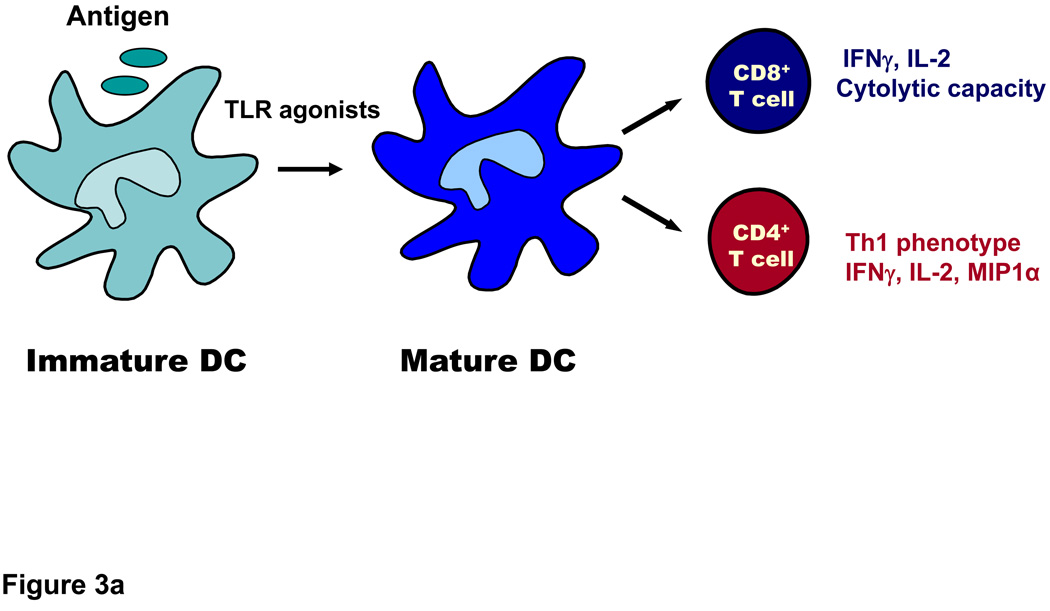
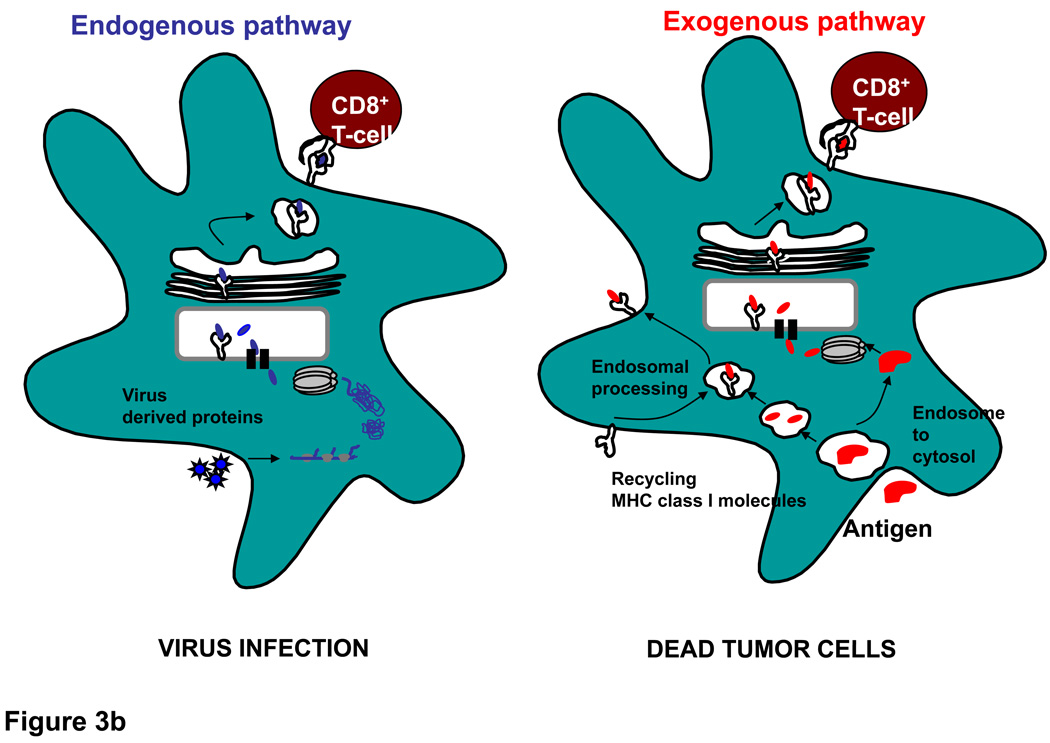
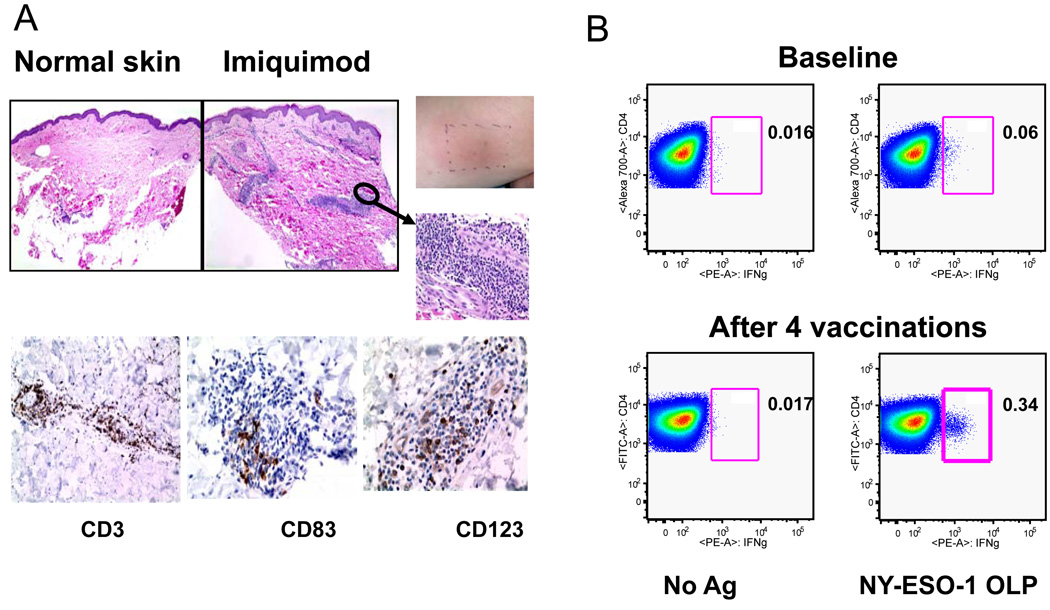
References
-
- Smyth MJ, Dunn GP, Schreiber RD. Cancer immunosurveillance and immunoediting: the roles of immunity in suppressing tumor development and shaping tumor immunogenicity. Advances in immunology. 2006;90:1–50. - PubMed
-
- Anderson KV, Bokla L, Nusslein-Volhard C. Establishment of dorsal-ventral polarity in the Drosophila embryo: the induction of polarity by the Toll gene product. Cell. 1985;42:791–798. - PubMed
-
- Pichlmair A, Reis e Sousa C. Innate recognition of viruses. Immunity. 2007;27:370–383. - PubMed
-
- Kawai T, Akira S. Innate immune recognition of viral infection. Nature immunology. 2006;7:131–137. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
