

Zinc availability regulates exit from meiosis in maturing mammalian oocytes
- PMID: 20693991
- PMCID: PMC2924620
- DOI: 10.1038/nchembio.419
Zinc availability regulates exit from meiosis in maturing mammalian oocytes
Abstract
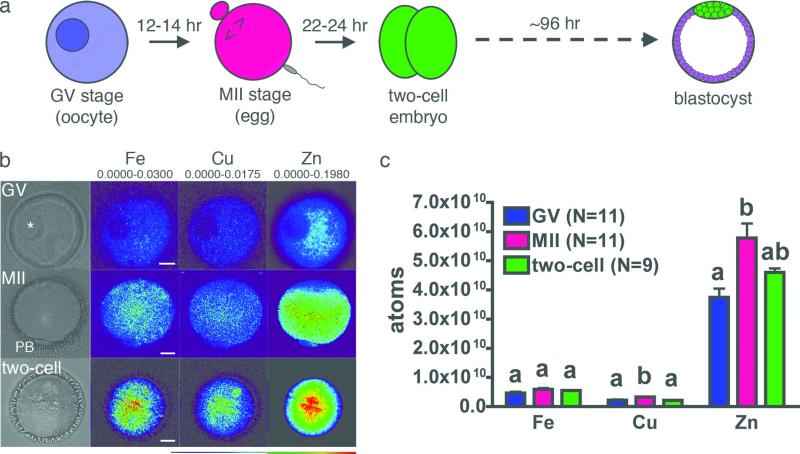
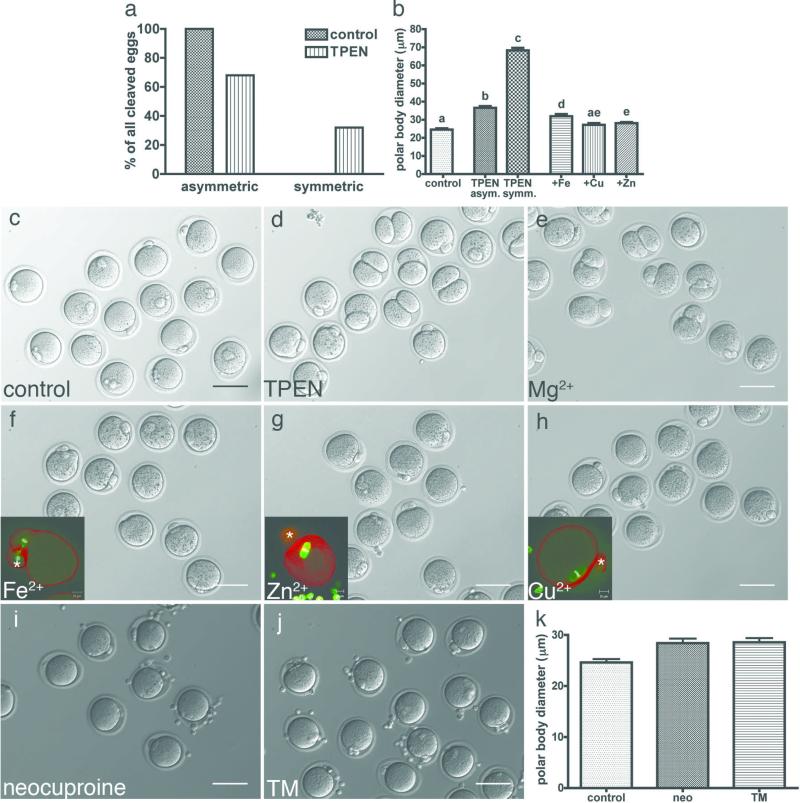
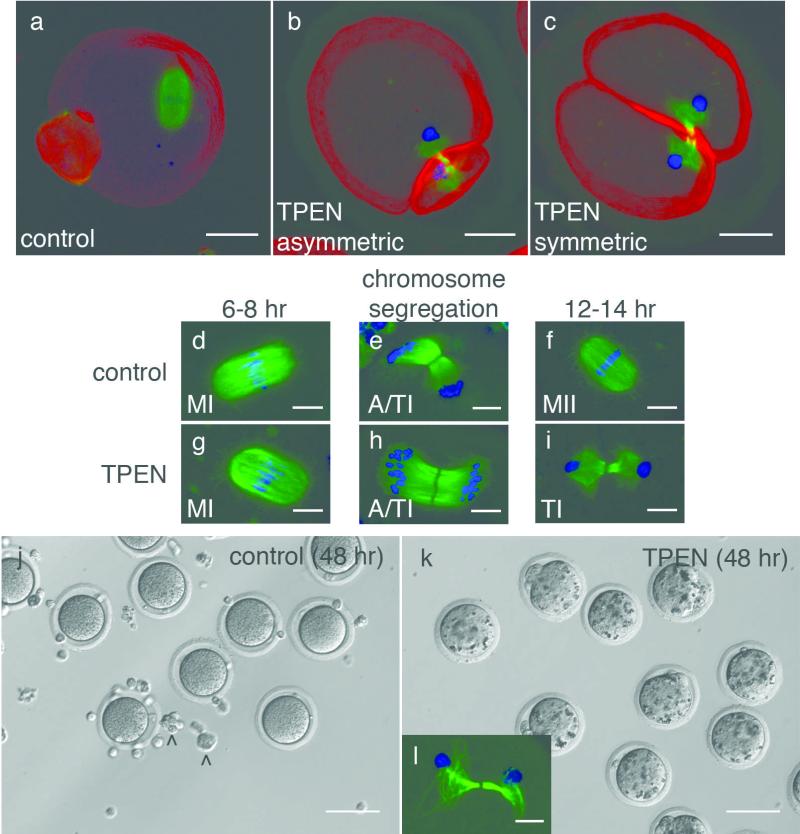
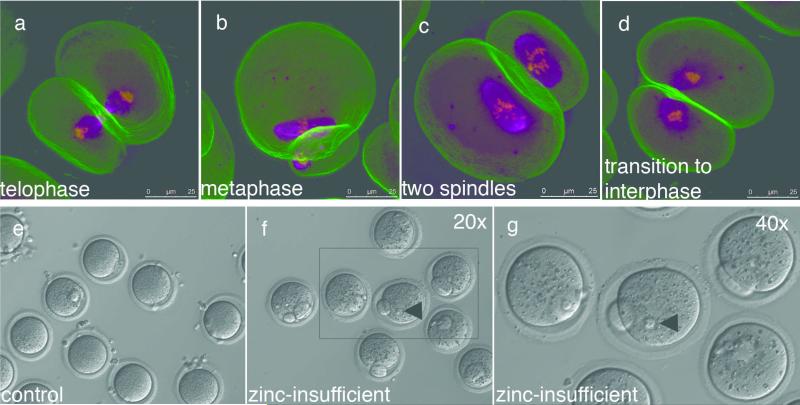
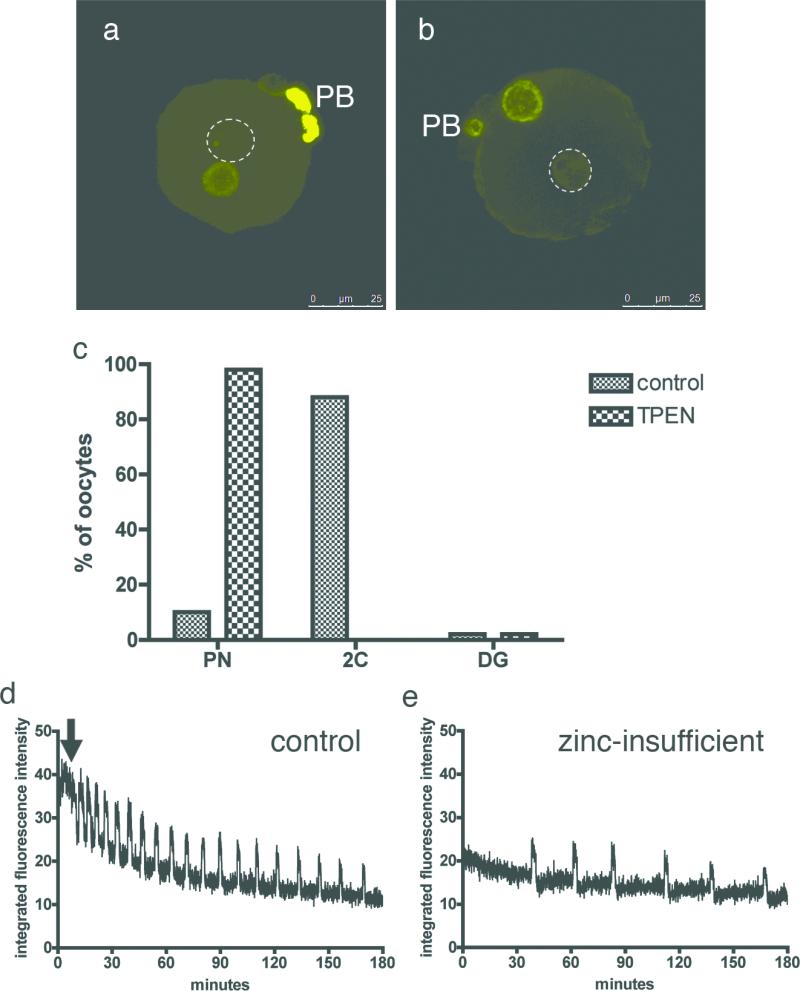
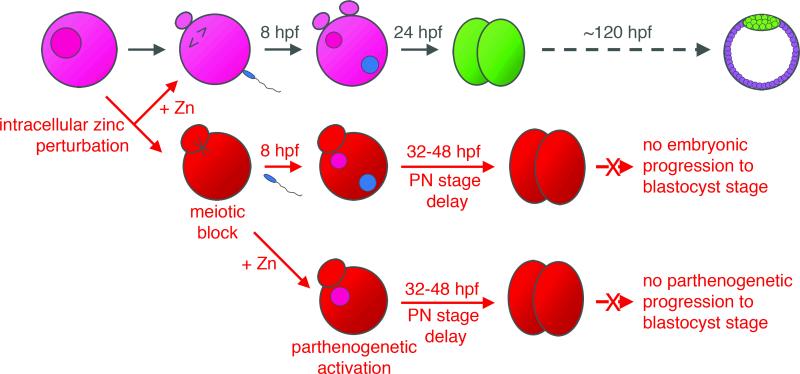
Cellular metal ion fluxes are known in alkali and alkaline earth metals but are not well documented in transition metals. Here we describe major changes in the zinc physiology of the mammalian oocyte as it matures and initiates embryonic development. Single-cell elemental analysis of mouse oocytes by synchrotron-based X-ray fluorescence microscopy (XFM) revealed a 50% increase in total zinc content within the 12-14-h period of meiotic maturation. Perturbation of zinc homeostasis with a cell-permeable small-molecule chelator blocked meiotic progression past telophase I. Zinc supplementation rescued this phenotype when administered before this meiotic block. However, after telophase arrest, zinc triggered parthenogenesis, suggesting that exit from this meiotic step is tightly regulated by the availability of a zinc-dependent signal. These results implicate the zinc bolus acquired during meiotic maturation as an important part of the maternal legacy to the embryo.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
