

Multiple distinct small RNAs originate from the same microRNA precursors
- PMID: 20696037
- PMCID: PMC2945783
- DOI: 10.1186/gb-2010-11-8-r81
Multiple distinct small RNAs originate from the same microRNA precursors
Abstract
Background: MicroRNAs (miRNAs), which originate from precursor transcripts with stem-loop structures, are essential gene expression regulators in eukaryotes.
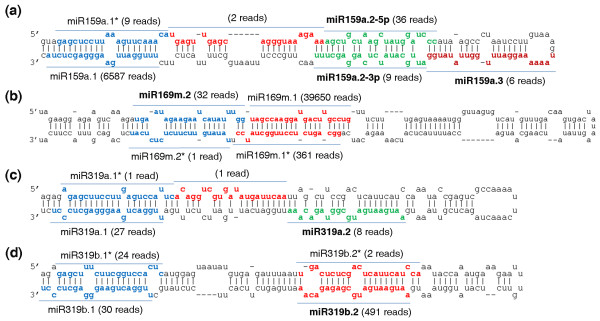
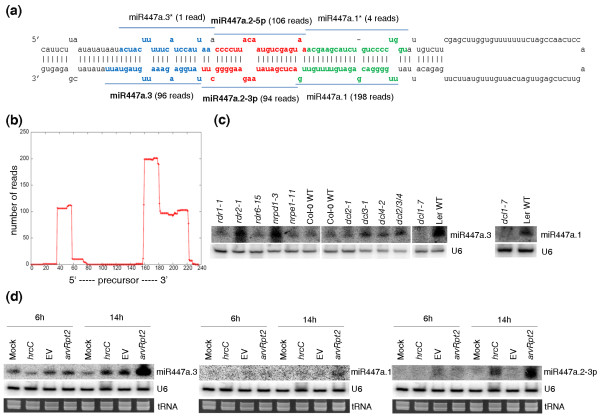
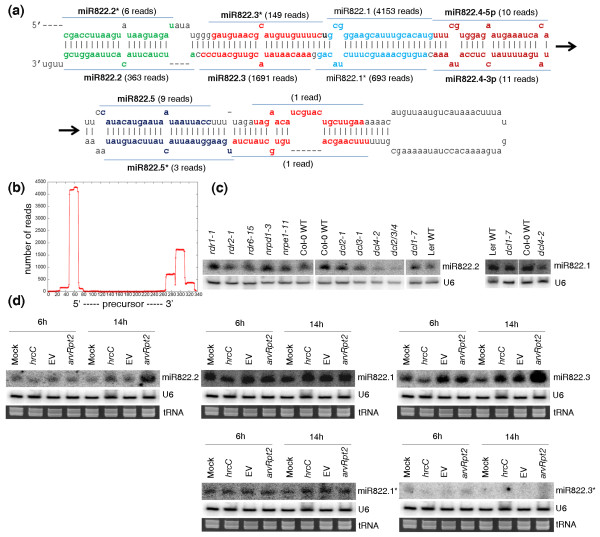
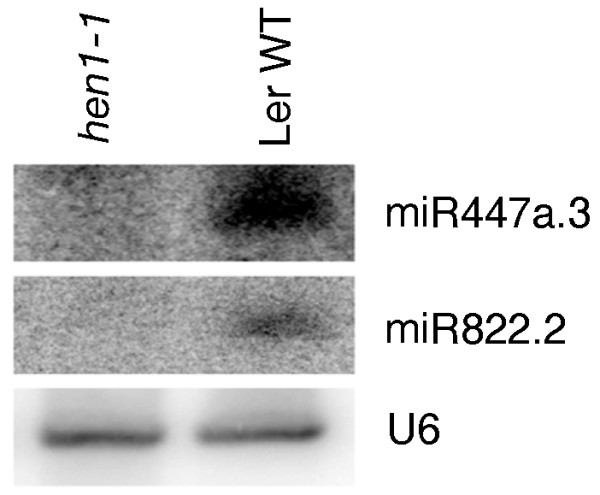
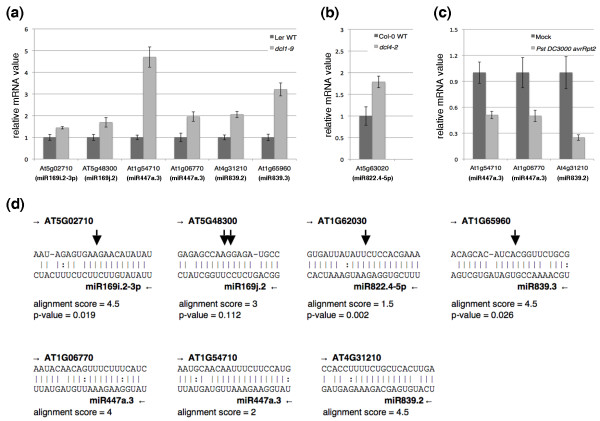
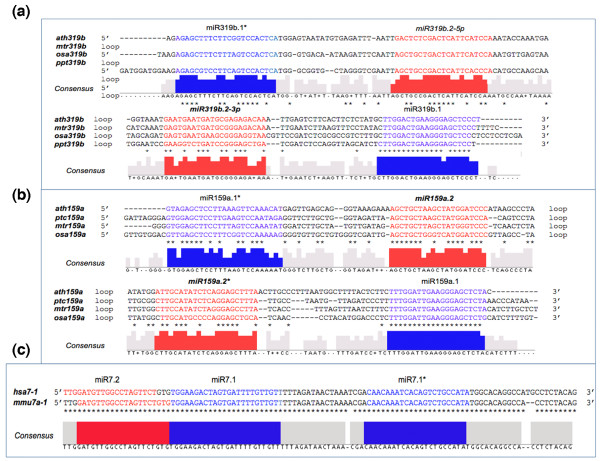
Results: We report 19 miRNA precursors in Arabidopsis that can yield multiple distinct miRNA-like RNAs in addition to miRNAs and miRNA*s. These miRNA precursor-derived miRNA-like RNAs are often arranged in phase and form duplexes with an approximately two-nucleotide 3'-end overhang. Their production depends on the same biogenesis pathway as their sibling miRNAs and does not require RNA-dependent RNA polymerases or RNA polymerase IV. These miRNA-like RNAs are methylated, and many of them are associated with Argonaute proteins. Some of the miRNA-like RNAs are differentially expressed in response to bacterial challenges, and some are more abundant than the cognate miRNAs. Computational and expression analyses demonstrate that some of these miRNA-like RNAs are potentially functional and they target protein-coding genes for silencing. The function of some of these miRNA-like RNAs was further supported by their target cleavage products from the published small RNA degradome data. Our systematic examination of public small-RNA deep sequencing data from four additional plant species (Oryza sativa, Physcomitrella patens, Medicago truncatula and Populus trichocarpa) and four animals (Homo sapiens, Mus musculus, Caenorhabditis elegans and Drosophila) shows that such miRNA-like RNAs exist broadly in eukaryotes.
Conclusions: We demonstrate that multiple miRNAs could derive from miRNA precursors by sequential processing of Dicer or Dicer-like proteins. Our results suggest that the pool of miRNAs is larger than was previously recognized, and miRNA-mediated gene regulation may be broader and more complex than previously thought.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
