

Maladaptive striatal plasticity in L-DOPA-induced dyskinesia
- PMID: 20696322
- PMCID: PMC2930606
- DOI: 10.1016/S0079-6123(10)83011-0
Maladaptive striatal plasticity in L-DOPA-induced dyskinesia
Abstract
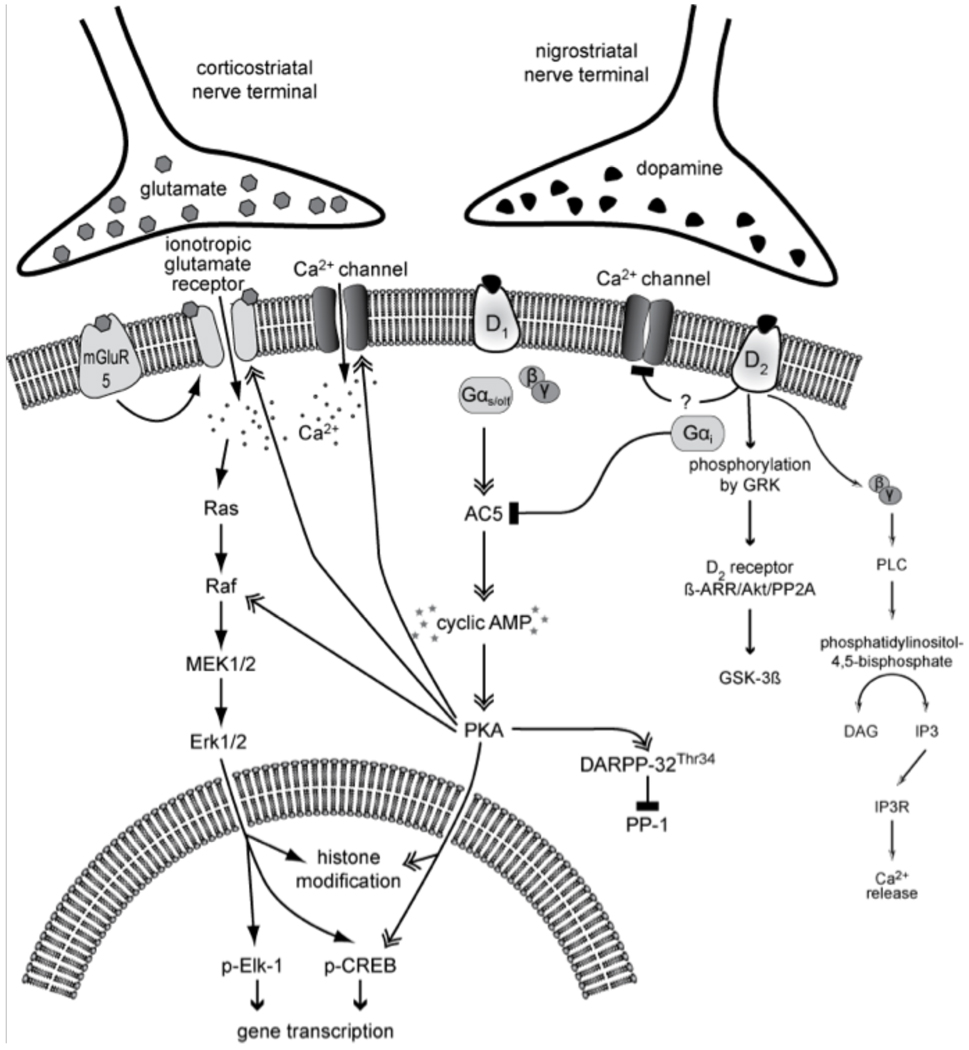
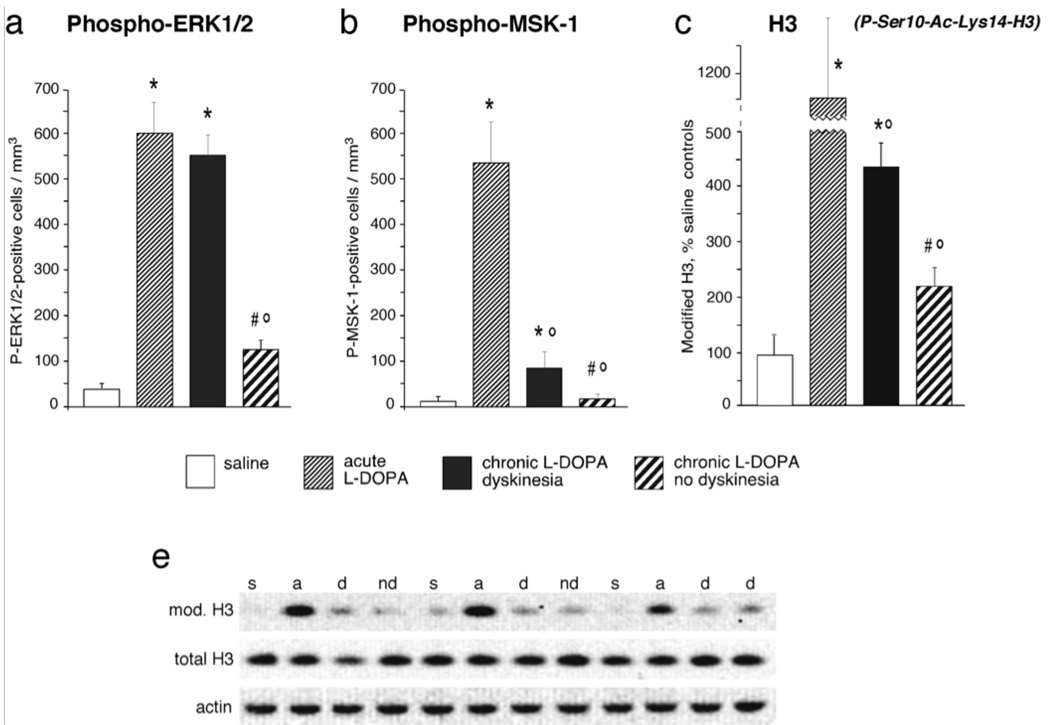
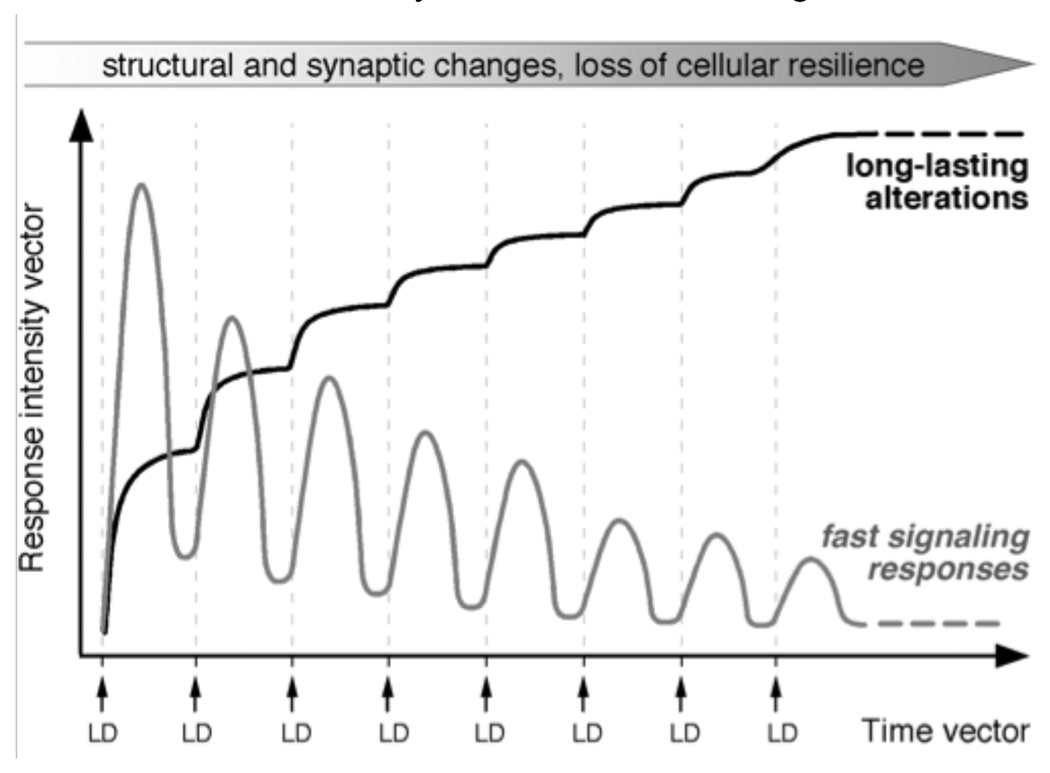
Dopamine (DA) replacement therapy with l-DOPA remains the most effective treatment for Parkinson's disease, but causes dyskinesia (abnormal involuntary movements) in the vast majority of the patients. The basic mechanisms of l-DOPA-induced dyskinesia (LID) have become the object of intense research focusing on neurochemical and molecular adaptations in the striatum. Here we review this vast literature and highlight trends that converge into a unifying pathophysiological interpretation. We propose that the core molecular alteration of striatal neurons in LID consists in an inability to turn down supersensitive signaling responses downstream of DA D1 receptors (where supersensitivity is primarily caused by DA denervation). The sustained activation of intracellular signaling pathways induced by each dose of l-DOPA leads to abnormal cellular plasticity and high bioenergetic expenditure. The over-exploitation of signaling pathways and energy reserves during treatment impairs the ability of striatal neurons to dynamically gate cortically driven motor commands. LID thus exemplifies a disorder where 'too much' molecular plasticity leads to plasticity failure in the striatum.
2010 Elsevier B.V. All rights reserved.
Figures

References
-
- Alonso-Frech F, Zamarbide I, Alegre M, Rodríguez-Oroz MC, Guridi J, Manrique M, Valencia M, Artieda J, Obeso JA. Slow oscillatory activity and levodopa-induced dyskinesias in Parkinson's disease. Brain. 2006;129(Pt7):1748–1757. - PubMed
-
- Andersson M, Hilbertson A, Cenci MA. Striatal fosB expression is causally linked with l-DOPA-induced abnormal involuntary movements and the associated upregulation of striatal prodynorphin mRNA in a rat model of Parkinson's disease. Neurobiol Dis. 1999;6(6):461–474. - PubMed
-
- Andersson M, Westin JE, Cenci MA. Time course of striatal DeltaFosB-like immunoreactivity and prodynorphin mRNA levels after discontinuation of chronic dopaminomimetic treatment. Eur J Neurosci. 2003;17(3):661–666. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
