

Molecular basis for the association of human E4B U box ubiquitin ligase with E2-conjugating enzymes UbcH5c and Ubc4
- PMID: 20696396
- PMCID: PMC3005147
- DOI: 10.1016/j.str.2010.04.017
Molecular basis for the association of human E4B U box ubiquitin ligase with E2-conjugating enzymes UbcH5c and Ubc4
Abstract
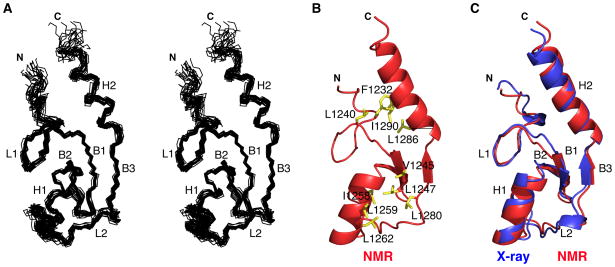
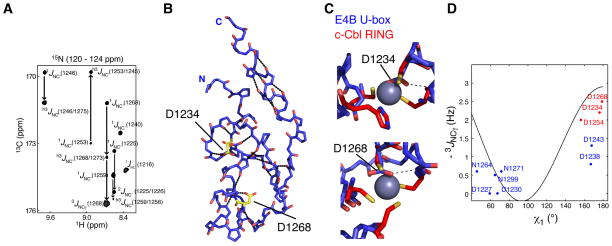
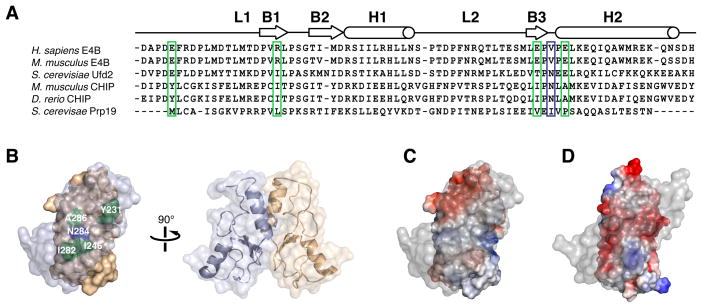
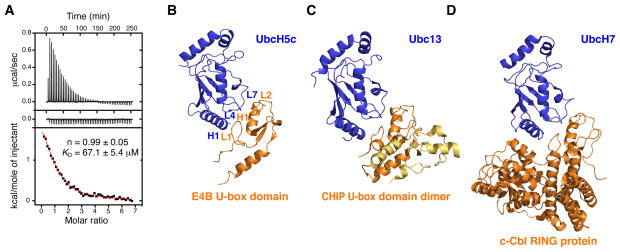
Human E4B, also called UFD2a, is a U box-containing protein that functions as an E3 ubiquitin ligase and an E4 polyubiquitin chain elongation factor. E4B is thought to participate in the proteasomal degradation of misfolded or damaged proteins through association with chaperones. The U box domain is an anchor site for E2 ubiquitin-conjugating enzymes, but little is known of the binding mechanism. Using X-ray crystallography and NMR spectroscopy, we determined the structures of E4B U box free and bound to UbcH5c and Ubc4 E2s. Whereas previously characterized U box domains are homodimeric, we show that E4B U box is a monomer stabilized by a network of hydrogen bonds identified from scalar coupling measurements. These structural studies, complemented by calorimetry- and NMR-based binding assays, suggest an allosteric regulation of UbcH5c and Ubc4 by E4B U box and provide a molecular basis to understand how the ubiquitylation machinery involving E4B assembles.
Copyright 2010 Elsevier Ltd. All rights reserved.
Figures



Comment in
-
Further insights into the ubiquitin pathway: understanding the scarlet letter code.Structure. 2010 Aug 11;18(8):891-2. doi: 10.1016/j.str.2010.07.004. Structure. 2010. PMID: 20696387
References
-
- Adams PD, Grosse-Kunstleve RW, Hung LW, Ioerger TR, McCoy AJ, Moriarty NW, Read RJ, Sacchettini JC, Sauter NK, Terwilliger TC. PHENIX: building new software for automated crystallographic structure determination. Acta Crystallogr Sect D. 2002;58:1948–1954. - PubMed
-
- Andersen P, Kragelund BB, Olsen AN, Larsen FH, Chua NH, Poulsen FM, Skriver K. Structure and biochemical function of a prototypical Arabidopsis U-box domain. J Biol Chem. 2004;279:40053–40061. - PubMed
-
- Aravind L, Koonin EV. The U box is a modified RING finger - a common domain in ubiquitination. Curr Biol. 2000;10:132–134. - PubMed
-
- Brzovic PS, Lissounov A, Christensen DE, Hoyt DW, Klevit RE. A UbcH5/ubiquitin noncovalent complex is required for processive BRCA1-directed ubiquitination. Mol Cell. 2006;21:873–880. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
