

KIF17 stabilizes microtubules and contributes to epithelial morphogenesis by acting at MT plus ends with EB1 and APC
- PMID: 20696710
- PMCID: PMC2922650
- DOI: 10.1083/jcb.201006044
KIF17 stabilizes microtubules and contributes to epithelial morphogenesis by acting at MT plus ends with EB1 and APC
Abstract
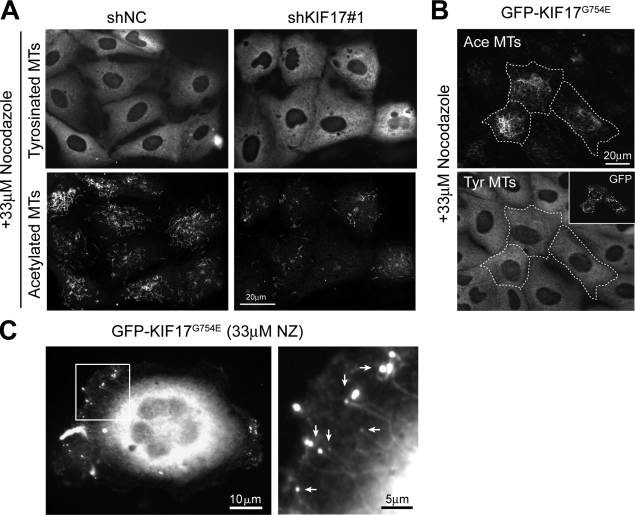
Epithelial polarization is associated with selective stabilization and reorganization of microtubule (MT) arrays. However, upstream events and downstream consequences of MT stabilization during epithelial morphogenesis are still unclear. We show that the anterograde kinesin KIF17 localizes to MT plus ends, stabilizes MTs, and affects epithelial architecture. Targeting of KIF17 to plus ends of growing MTs requires kinesin motor activity and interaction with EB1. In turn, KIF17 participates in localizing adenomatous polyposis coli (APC) to the plus ends of a subset of MTs. We found that KIF17 affects MT dynamics, polymerization rates, and MT plus end stabilization to generate posttranslationally acetylated MTs. Depletion of KIF17 from cells growing in three-dimensional matrices results in aberrant epithelial cysts that fail to generate a single central lumen and to polarize apical markers. These findings implicate KIF17 in MT stabilization events that contribute to epithelial polarization and morphogenesis.
Figures









Similar articles
-
Direct regulation of microtubule dynamics by KIF17 motor and tail domains.J Biol Chem. 2013 Nov 8;288(45):32302-32313. doi: 10.1074/jbc.M113.494989. Epub 2013 Sep 26. J Biol Chem. 2013. PMID: 24072717 Free PMC article.
-
Adenomatous polyposis coli on microtubule plus ends in cell extensions can promote microtubule net growth with or without EB1.Mol Biol Cell. 2006 May;17(5):2331-45. doi: 10.1091/mbc.e05-06-0498. Epub 2006 Mar 8. Mol Biol Cell. 2006. PMID: 16525027 Free PMC article.
-
A biosensor of local kinesin activity reveals roles of PKC and EB1 in KIF17 activation.J Cell Biol. 2013 Nov 11;203(3):445-55. doi: 10.1083/jcb.201305023. Epub 2013 Nov 4. J Cell Biol. 2013. PMID: 24189273 Free PMC article.
-
TIP maker and TIP marker; EB1 as a master controller of microtubule plus ends.J Cell Biol. 2005 Oct 24;171(2):197-200. doi: 10.1083/jcb.200509150. J Cell Biol. 2005. PMID: 16247021 Free PMC article. Review.
-
HIV-1 capsid exploitation of the host microtubule cytoskeleton during early infection.Retrovirology. 2021 Jul 6;18(1):19. doi: 10.1186/s12977-021-00563-3. Retrovirology. 2021. PMID: 34229718 Free PMC article. Review.
Cited by
-
Role of membrane traffic in the generation of epithelial cell asymmetry.Nat Cell Biol. 2012 Dec;14(12):1235-43. doi: 10.1038/ncb2635. Nat Cell Biol. 2012. PMID: 23196841 Free PMC article. Review.
-
Polarized sorting and trafficking in epithelial cells.Cell Res. 2012 May;22(5):793-805. doi: 10.1038/cr.2012.64. Epub 2012 Apr 24. Cell Res. 2012. PMID: 22525333 Free PMC article. Review.
-
Ca(2+)/Calmodulin-Dependent Protein Kinase IV Promotes Interplay of Proteins in Chromatoid Body of Male Germ Cells.Sci Rep. 2015 Jul 16;5:12126. doi: 10.1038/srep12126. Sci Rep. 2015. PMID: 26179157 Free PMC article.
-
Kif4 interacts with EB1 and stabilizes microtubules downstream of Rho-mDia in migrating fibroblasts.PLoS One. 2014 Mar 21;9(3):e91568. doi: 10.1371/journal.pone.0091568. eCollection 2014. PLoS One. 2014. PMID: 24658398 Free PMC article.
-
Microtubule Motors in Establishment of Epithelial Cell Polarity.Cold Spring Harb Perspect Biol. 2018 Feb 1;10(2):a027896. doi: 10.1101/cshperspect.a027896. Cold Spring Harb Perspect Biol. 2018. PMID: 28264820 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
