

Vinca drug components accumulate exclusively in leaf exudates of Madagascar periwinkle
- PMID: 20696903
- PMCID: PMC2930567
- DOI: 10.1073/pnas.0911451107
Vinca drug components accumulate exclusively in leaf exudates of Madagascar periwinkle
Abstract
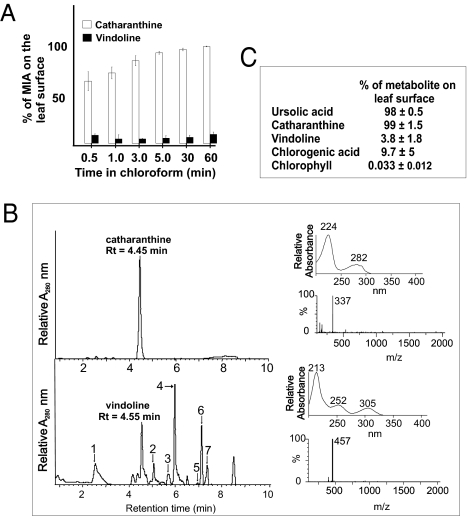
The monoterpenoid indole alkaloids (MIAs) of Madagascar periwinkle (Catharanthus roseus) continue to be the most important source of natural drugs in chemotherapy treatments for a range of human cancers. These anticancer drugs are derived from the coupling of catharanthine and vindoline to yield powerful dimeric MIAs that prevent cell division. However the precise mechanisms for their assembly within plants remain obscure. Here we report that the complex development-, environment-, organ-, and cell-specific controls involved in expression of MIA pathways are coupled to secretory mechanisms that keep catharanthine and vindoline separated from each other in living plants. Although the entire production of catharanthine and vindoline occurs in young developing leaves, catharanthine accumulates in leaf wax exudates of leaves, whereas vindoline is found within leaf cells. The spatial separation of these two MIAs provides a biological explanation for the low levels of dimeric anticancer drugs found in the plant that result in their high cost of commercial production. The ability of catharanthine to inhibit the growth of fungal zoospores at physiological concentrations found on the leaf surface of Catharanthus leaves, as well as its insect toxicity, provide an additional biological role for its secretion. We anticipate that this discovery will trigger a broad search for plants that secrete alkaloids, the biological mechanisms involved in their secretion to the plant surface, and the ecological roles played by them.
Conflict of interest statement
The authors declare no conflict of interest.
Figures


References
-
- Mangeney P, Andriamialisoa RZ, Langlois N, Langlois Y, Potier P. Preparation of vinblastine, vincristine, and leurosidine, antitumor alkaloids from Catharanthus species (Apocynaceae) J Am Chem Soc. 1979;101:2243–2245.
-
- Kutney JP, et al. A highly efficient and commercially important synthesis of the antitumor catharanthus alkaloids vinblastine and leurosidine from catharanthine and vindoline. Heterocycles. 1988;27:1845–1853.
-
- Kuehne ME, Matson PA, Bornmann WG. Enantioselective syntheses of vinblastine, leurosidine, vincovaline and 20'-epi-vincovaline. J Org Chem. 1991;56:513–528.
-
- Magnus P, Mendoza JS, Stamford A, Ladlow M, Willis P. Nonoxidative coupling methodology for the synthesis of the antitumor bisindole alkaloid vinblastine and a lower-half analog: Solvent effect on the stereochemistry of the crucial C-15/C-18' bond. J Am Chem Soc. 1992;114:10232–10245.
-
- Yokoshima S, et al. Stereocontrolled total synthesis of (+)-vinblastine. J Am Chem Soc. 2002;124:2137–2139. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
