

The reactive tumor microenvironment: MUC1 signaling directly reprograms transcription of CTGF
- PMID: 20697347
- PMCID: PMC3412169
- DOI: 10.1038/onc.2010.327
The reactive tumor microenvironment: MUC1 signaling directly reprograms transcription of CTGF
Abstract
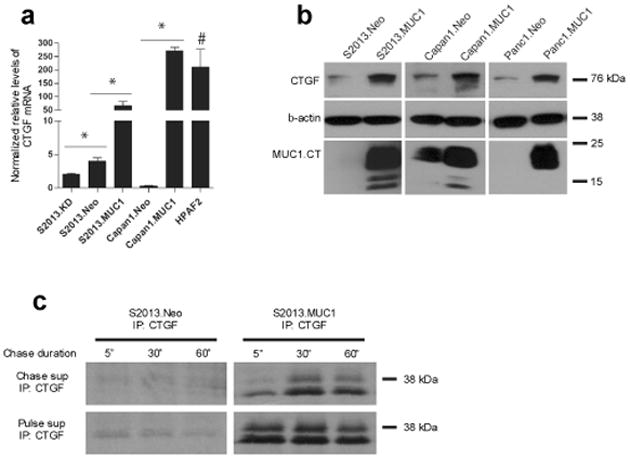
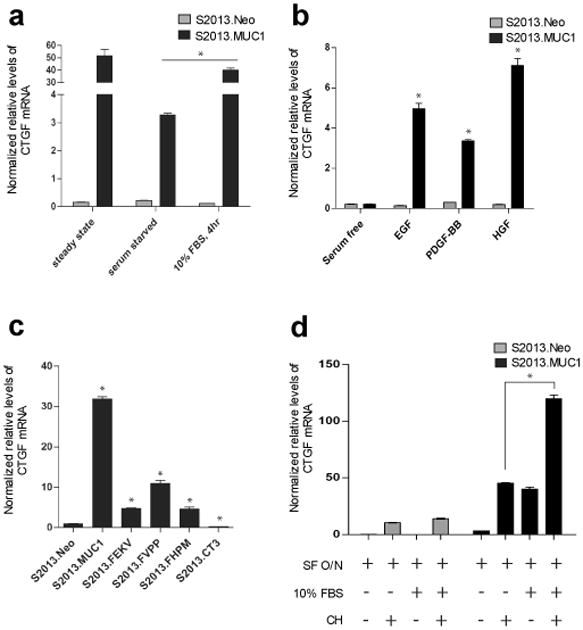
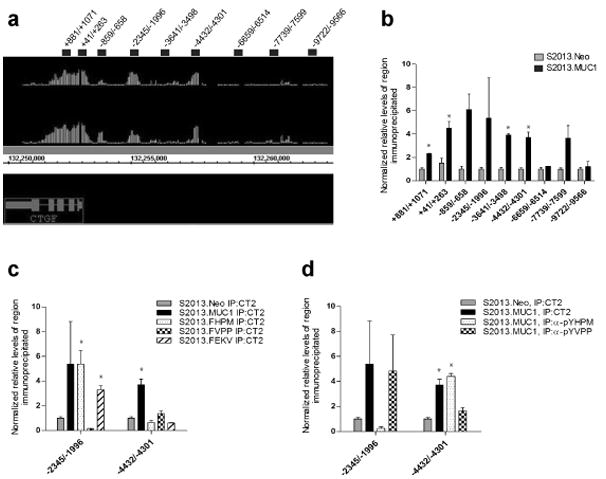
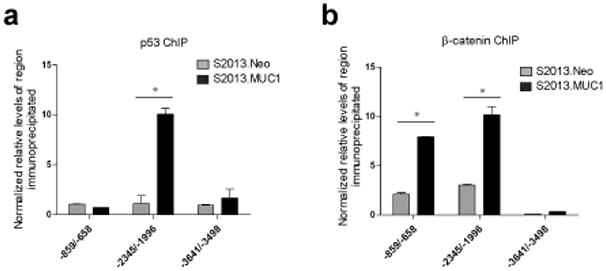
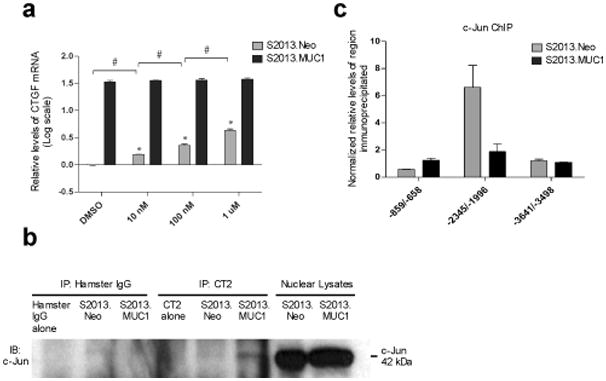
The MUC1 cytoplasmic tail (MUC1.CT) conducts signals from spatial and extracellular cues (growth factor and cytokine stimulation) to evoke a reprogramming of the cellular transcriptional profile. Specific phosphorylated forms of the MUC1.CT achieve this function by differentially associating with transcription factors and redirecting their transcriptional regulatory capabilities at specific gene regulatory elements. The specificity of interaction between MUC1.CT and several transcription factors is dictated by the phosphorylation pattern of the 18 potential phosphorylation motifs within the MUC1.CT. To better appreciate the scope of differential gene expression triggered by MUC1.CT activation, we performed microarray gene expression analysis and chromatin immunoprecipitation (ChIP)-chip promoter analysis and identified the genome-wide transcriptional targets of MUC1.CT signaling in pancreatic cancer. On a global scale, MUC1.CT preferentially targets genes related to invasion, angiogenesis and metastasis, suggesting that MUC1.CT signaling contributes to establishing a reactive tumor microenvironment during tumor progression to metastatic disease. We examined in detail the molecular mechanisms of MUC1.CT signaling that induces the expression of connective tissue growth factor (CTGF/CCN2), a potent mediator of ECM remodeling and angiogenesis. We demonstrate a robust induction of CTGF synthesis and secretion in response to serum factors that is enabled only when MUC1 is highly expressed. We demonstrate the requirement of phosphorylation at distinct tyrosine motifs within the MUC1.CT for MUC1-induced CTGF expression and demonstrate a phosphorylation-specific localization of MUC1.CT to the CTGF promoter. We found that MUC1 reorganizes transcription factor occupancy of genomic regions upstream of the CTGF gene, directing β-catenin and mutant p53 to CTGF gene regulatory elements to promote CTGF expression and destabilizing the interaction at these regions of the transcriptional repressor, c-Jun. With this example we illustrate the capacity of MUC1.CT to mediate transcription factor activity in a context-dependent manner to achieve wide spread and robust changes in gene expression and facilitate creation of the reactive tumor microenvironment.
Conflict of interest statement
Figures
Comment in
-
Oncogenic function of the MUC1 receptor subunit in gene regulation.Oncogene. 2010 Oct 21;29(42):5663-6. doi: 10.1038/onc.2010.334. Epub 2010 Aug 16. Oncogene. 2010. PMID: 20711235 Free PMC article.
References
-
- Aikawa T, Gunn J, Spong SM, Klaus SJ, Korc M. Connective tissue growth factor-specific antibody attenuates tumor growth, metastasis, and angiogenesis in an orthotopic mouse model of pancreatic cancer. Mol Cancer Ther. 2006;5:1108–1116. - PubMed
-
- Brigstock DR. The CCN family: A new stimulus package. J Endocrinol. 2003;178:169–175. - PubMed
-
- Burdick MD, Harris A, Reid CJ, Iwamura T, Hollingsworth MA. Oligosaccharides expressed on MUC1 produced by pancreatic and colon tumor cell lines. J Biol Chem. 1997;272:24198–24202. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
