

Characterization of surface-exposed reactive cysteine residues in Saccharomyces cerevisiae
- PMID: 20698499
- PMCID: PMC3061811
- DOI: 10.1021/bi100677a
Characterization of surface-exposed reactive cysteine residues in Saccharomyces cerevisiae
Abstract
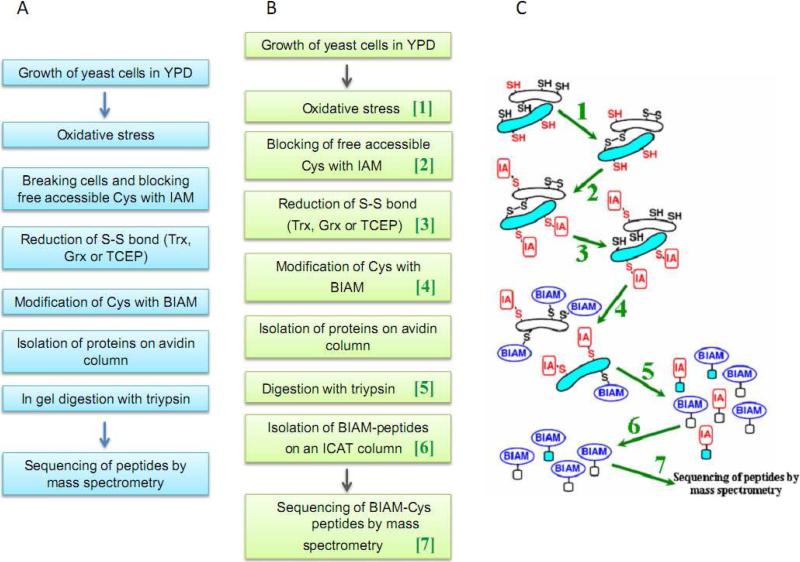
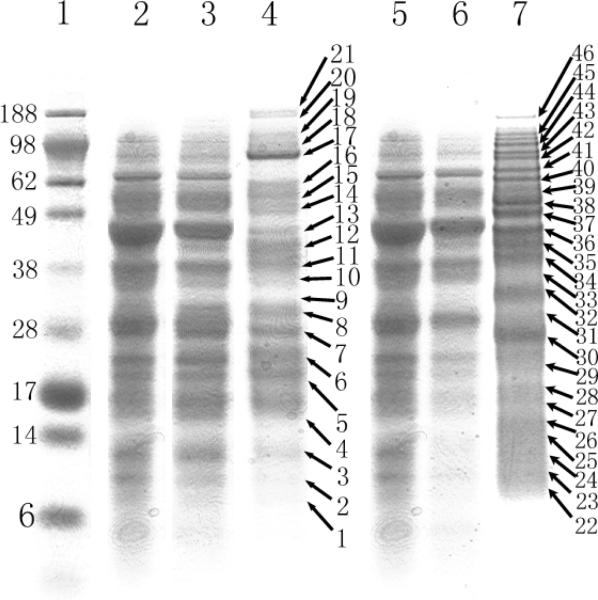
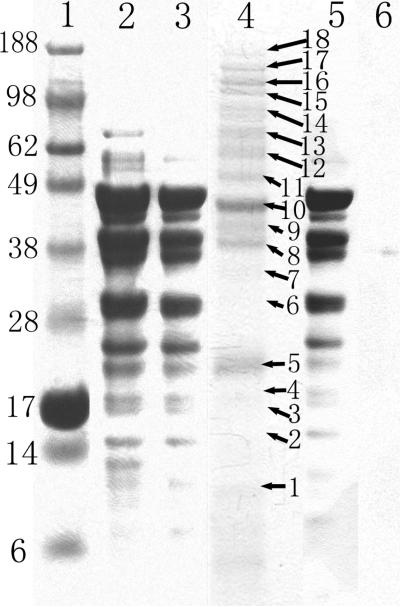
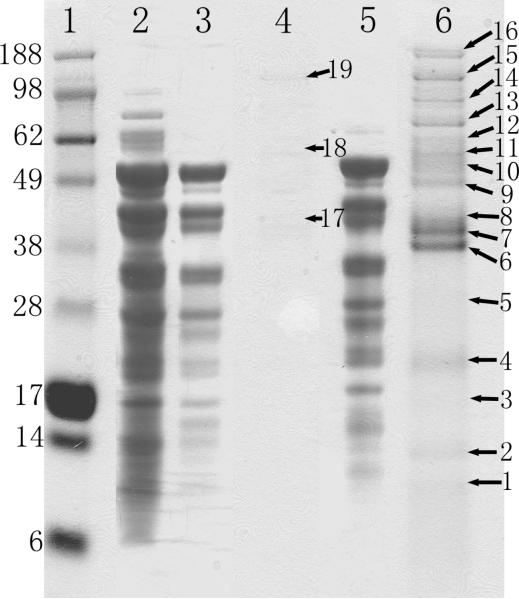
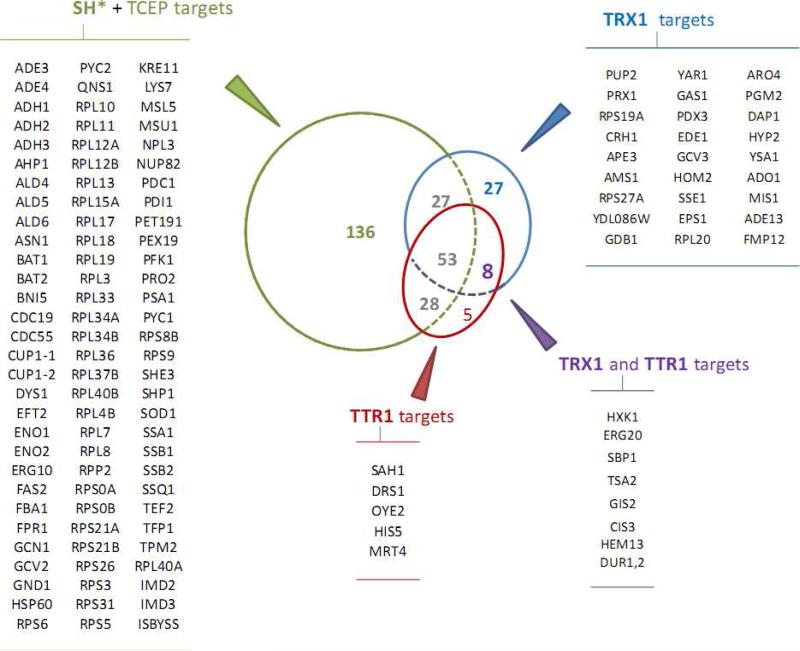
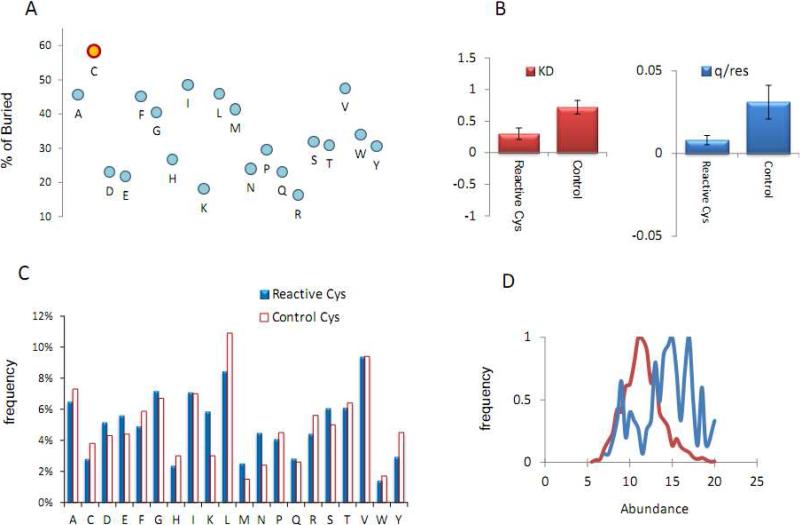
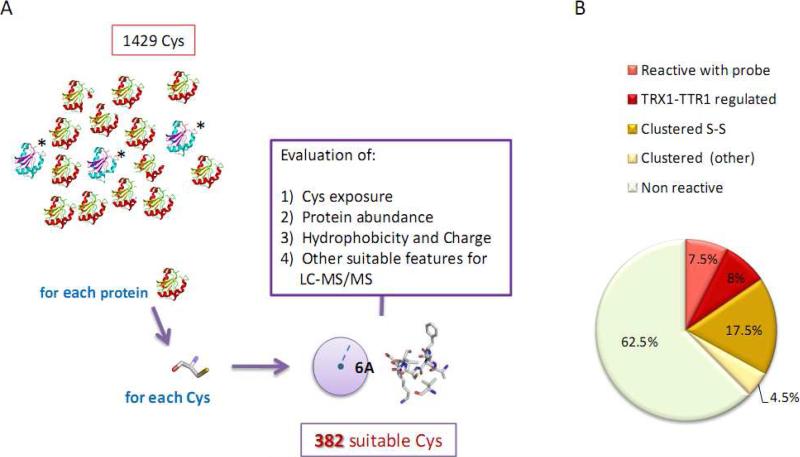
Numerous cellular processes are subject to redox regulation, and thiol-dependent redox control, acting through reactive cysteine (Cys) residues, is among the major mechanisms of redox regulation. However, information on the sets of proteins that provide thiol-based redox regulation or are affected by it is limited. Here, we describe proteomic approaches to characterize proteins that contain reactive thiols and methods to identify redox Cys in these proteins. Using Saccharomyces cerevisiae as a eukaryotic model organism, we identified 284 proteins with exposed reactive Cys and determined the identities of 185 of these residues. We then characterized subsets of these proteins as in vitro targets of major cellular thiol oxidoreductases, thioredoxin and glutaredoxin, and found that these enzymes can control the redox state of a significant number of thiols in target proteins. We further examined common features of exposed reactive Cys and compared them with an unbiased control set of Cys using computational approaches. This analysis (i) validated the efficacy of targeting exposed Cys in proteins in their native, folded state, (ii) quantified the proportion of targets that can be redox regulated via thiol oxidoreductase systems, and (iii) revealed the theoretical range of the experimental approach with regard to protein abundance and physicochemical properties of reactive Cys. From these analyses, we estimate that approximately one-fourth of exposed Cys in the yeast proteome can be regarded as functional sites, either subject to regulation by thiol oxidoreductases or involved in structural disulfides and metal binding.
Figures
Similar articles
-
A structure-based approach for detection of thiol oxidoreductases and their catalytic redox-active cysteine residues.PLoS Comput Biol. 2009 May;5(5):e1000383. doi: 10.1371/journal.pcbi.1000383. Epub 2009 May 8. PLoS Comput Biol. 2009. PMID: 19424433 Free PMC article.
-
Thiol redox proteomics identifies differential targets of cytosolic and mitochondrial glutaredoxin-2 isoforms in Saccharomyces cerevisiae. Reversible S-glutathionylation of DHBP synthase (RIB3).J Proteomics. 2011 Oct 19;74(11):2487-97. doi: 10.1016/j.jprot.2011.04.018. Epub 2011 Apr 30. J Proteomics. 2011. PMID: 21565288
-
Redox potentials of active-site bis(cysteinyl) fragments of thiol-protein oxidoreductases.Biochemistry. 1993 Jul 27;32(29):7488-95. doi: 10.1021/bi00080a021. Biochemistry. 1993. PMID: 8338847
-
Thiol redox homeostasis in neurodegenerative disease.Redox Biol. 2015 Aug;5:186-194. doi: 10.1016/j.redox.2015.04.004. Epub 2015 Apr 22. Redox Biol. 2015. PMID: 25974624 Free PMC article. Review.
-
[Characterization of some thiol oxidoreductase family members].Mol Biol (Mosk). 2013 Jul-Aug;47(4):568-82. doi: 10.7868/s0026898413040149. Mol Biol (Mosk). 2013. PMID: 24466746 Review. Russian.
Cited by
-
Global analysis of protein aggregation in yeast during physiological conditions and arsenite stress.Biol Open. 2014 Sep 12;3(10):913-23. doi: 10.1242/bio.20148938. Biol Open. 2014. PMID: 25217615 Free PMC article.
-
Multimolecular salivary mucin complex is altered in saliva of cigarette smokers: detection of disulfide bridges by Raman spectroscopy.Biomed Res Int. 2013;2013:168765. doi: 10.1155/2013/168765. Epub 2012 Dec 26. Biomed Res Int. 2013. PMID: 23509686 Free PMC article.
-
Distinguishable short-term effects of tea and water drinking on human saliva redox.NPJ Sci Food. 2024 Apr 22;8(1):22. doi: 10.1038/s41538-024-00266-x. NPJ Sci Food. 2024. PMID: 38649360 Free PMC article.
-
Cysteine-mediated redox signaling: chemistry, biology, and tools for discovery.Chem Rev. 2013 Jul 10;113(7):4633-79. doi: 10.1021/cr300163e. Epub 2013 Mar 20. Chem Rev. 2013. PMID: 23514336 Free PMC article. Review. No abstract available.
-
Redox regulation of proteostasis.J Biol Chem. 2024 Dec;300(12):107977. doi: 10.1016/j.jbc.2024.107977. Epub 2024 Nov 8. J Biol Chem. 2024. PMID: 39522946 Free PMC article. Review.
References
-
- Stadtman ER. Oxidation of free amino acids and amino acid residues in proteins by radiolysis and by metal-catalyzed reactions. Annu. Rev. Biochem. 1993;62:797–821. - PubMed
-
- Stadtman ER, Levine RL. Free radical-mediated oxidation of free amino acids and amino acid residues in proteins. Amino Acids. 2003;25:207–218. - PubMed
-
- Berlett BS, Stadtman ER. Protein oxidation in aging, disease, and oxidative stress. J. Biol. Chem. 1997;272:20313–20316. - PubMed
-
- Poole LB, Karplus PA, Claiborne A. Protein sulfenic acids in redox signaling. Annu Rev Pharmacol Toxico. 2004;44:325–347. - PubMed
-
- Barford D. The role of cysteine residues as redox-sensitive regulatory switches. Curr Opin Struct Biol. 2004;14:679–686. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
