

Lamellipodium extension and membrane ruffling require different SNARE-mediated trafficking pathways
- PMID: 20698987
- PMCID: PMC2925818
- DOI: 10.1186/1471-2121-11-62
Lamellipodium extension and membrane ruffling require different SNARE-mediated trafficking pathways
Abstract
Background: Intracellular membrane traffic is an essential component of the membrane remodeling that supports lamellipodium extension during cell adhesion. The membrane trafficking pathways that contribute to cell adhesion have not been fully elucidated, but recent studies have implicated SNARE proteins. Here, the functions of several SNAREs (SNAP23, VAMP3, VAMP4 and syntaxin13) are characterized during the processes of cell spreading and membrane ruffling.
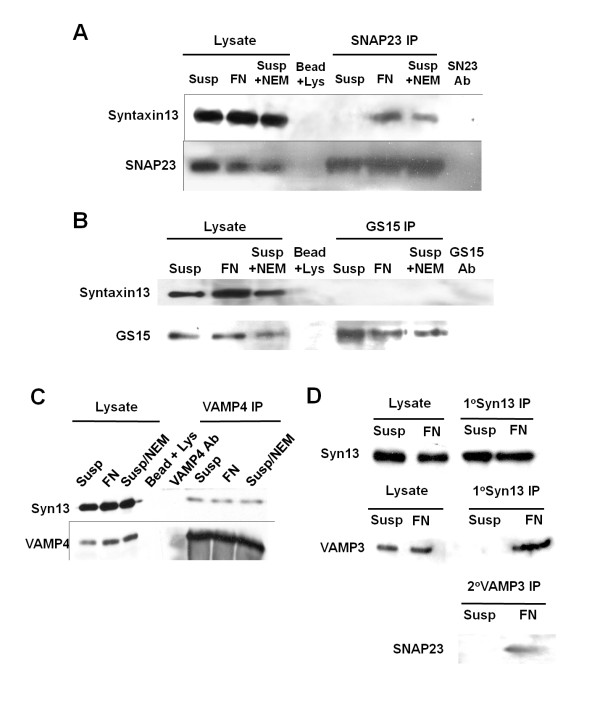
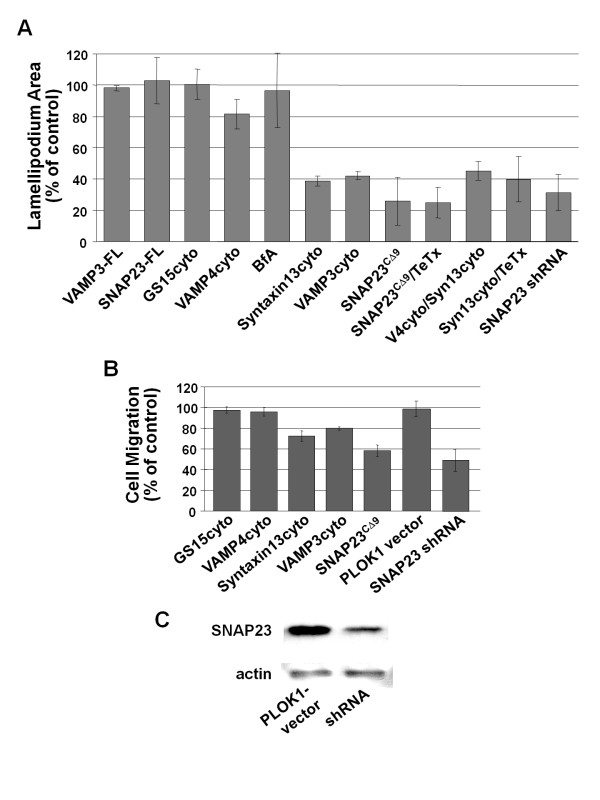
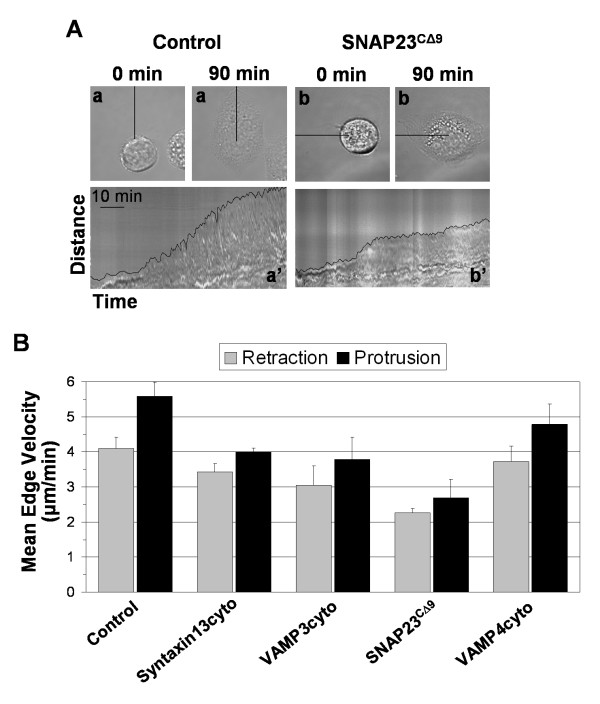
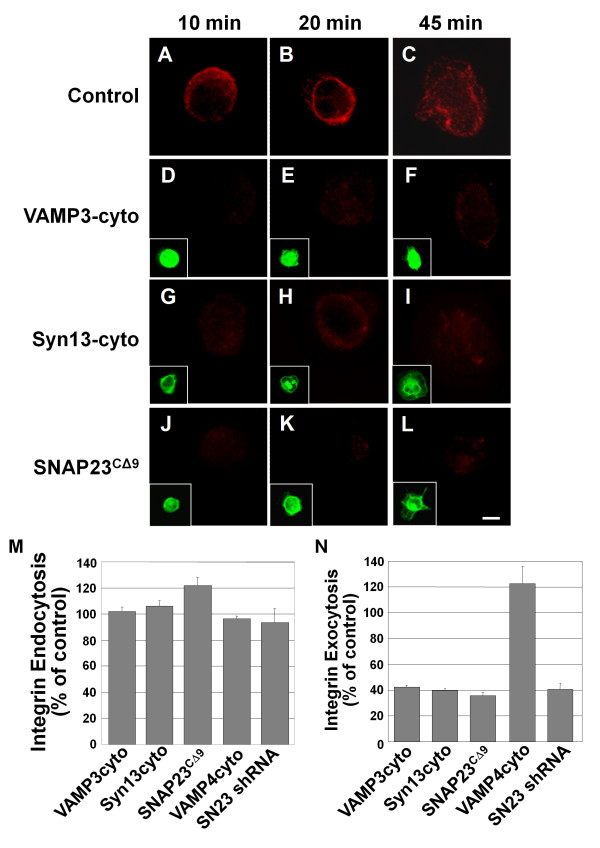
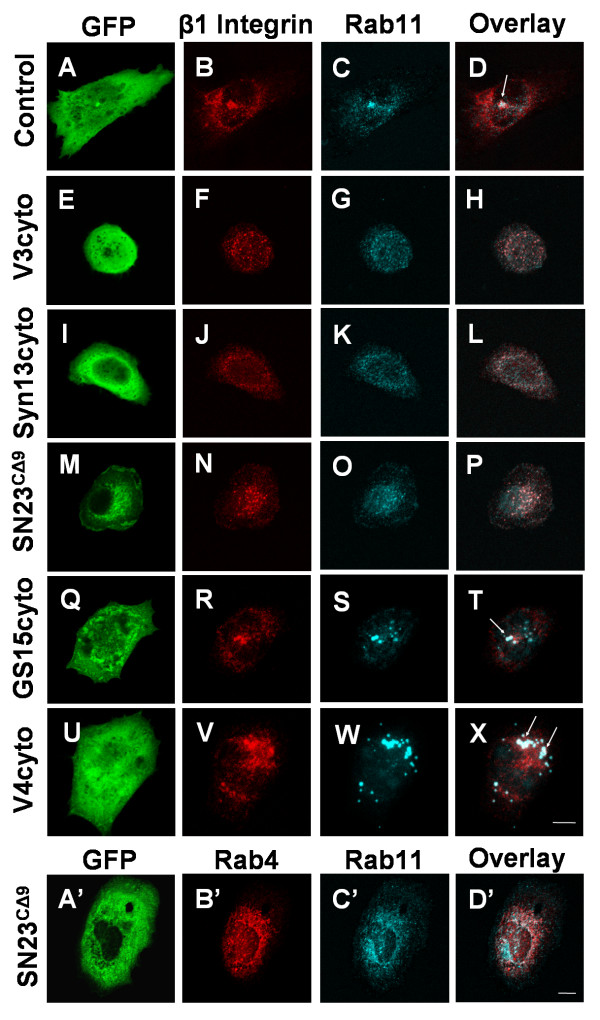
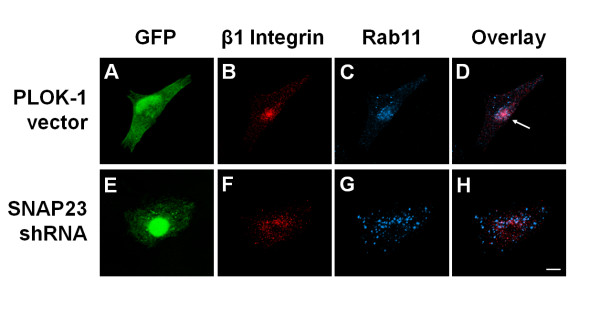
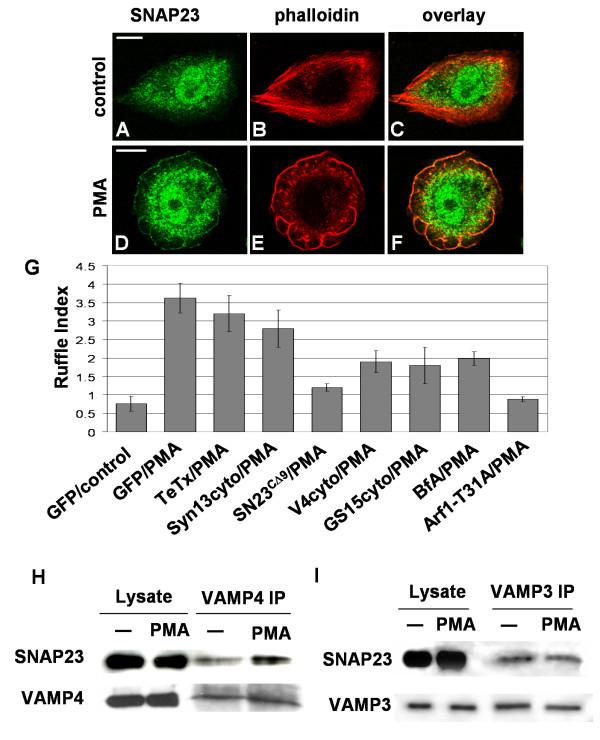
Results: We report the first description of a SNARE complex, containing SNAP23, syntaxin13 and cellubrevin/VAMP3, that is induced by cell adhesion to an extracellular matrix. Impairing the function of the SNAREs in the complex using inhibitory SNARE domains disrupted the recycling endosome, impeded delivery of integrins to the cell surface, and reduced haptotactic cell migration and spreading. Blocking SNAP23 also inhibited the formation of PMA-stimulated, F-actin-rich membrane ruffles; however, membrane ruffle formation was not significantly altered by inhibition of VAMP3 or syntaxin13. In contrast, membrane ruffling, and not cell spreading, was sensitive to inhibition of two SNAREs within the biosynthetic secretory pathway, GS15 and VAMP4. Consistent with this, formation of a complex containing VAMP4 and SNAP23 was enhanced by treatment of cells with PMA. The results reveal a requirement for the function of a SNAP23-syntaxin13-VAMP3 complex in the formation of lamellipodia during cell adhesion and of a VAMP4-SNAP23-containing complex during PMA-induced membrane ruffling.
Conclusions: Our findings suggest that different SNARE-mediated trafficking pathways support membrane remodeling during ECM-induced lamellipodium extension and PMA-induced ruffle formation, pointing to important mechanistic differences between these processes.
Figures
Similar articles
-
Recycling endosome membrane incorporation into the leading edge regulates lamellipodia formation and macrophage migration.Traffic. 2010 Oct;11(10):1370-9. doi: 10.1111/j.1600-0854.2010.01094.x. Traffic. 2010. PMID: 20604897
-
VAMP3, syntaxin-13 and SNAP23 are involved in secretion of matrix metalloproteinases, degradation of the extracellular matrix and cell invasion.J Cell Sci. 2009 Nov 15;122(Pt 22):4089-98. doi: 10.1242/jcs.052761. J Cell Sci. 2009. PMID: 19910495
-
SNARE-mediated trafficking of alpha5beta1 integrin is required for spreading in CHO cells.Biochem Biophys Res Commun. 2005 Oct 7;335(4):1199-210. doi: 10.1016/j.bbrc.2005.07.195. Biochem Biophys Res Commun. 2005. PMID: 16112083
-
Role of cholesterol in SNARE-mediated trafficking on intracellular membranes.J Cell Sci. 2015 Mar 15;128(6):1071-81. doi: 10.1242/jcs.164459. Epub 2015 Feb 4. J Cell Sci. 2015. PMID: 25653390 Review.
-
The cystic fibrosis transmembrane conductance regulator's expanding SNARE interactome.Traffic. 2011 Apr;12(4):364-71. doi: 10.1111/j.1600-0854.2011.01161.x. Epub 2011 Feb 8. Traffic. 2011. PMID: 21223469 Review.
Cited by
-
Endothelial cell migration on fibronectin is regulated by syntaxin 6-mediated alpha5beta1 integrin recycling.J Biol Chem. 2011 Oct 21;286(42):36749-61. doi: 10.1074/jbc.M111.260828. Epub 2011 Aug 31. J Biol Chem. 2011. PMID: 21880737 Free PMC article.
-
Radial Migration Dynamics Is Modulated in a Laminar and Area-Specific Manner During Primate Corticogenesis.Front Cell Dev Biol. 2020 Oct 16;8:588814. doi: 10.3389/fcell.2020.588814. eCollection 2020. Front Cell Dev Biol. 2020. PMID: 33178700 Free PMC article.
-
Autophagy-independent role of ATG9A vesicles as carriers for galectin-9 secretion.Nat Commun. 2025 May 7;16(1):4259. doi: 10.1038/s41467-025-59605-5. Nat Commun. 2025. PMID: 40335523 Free PMC article.
-
Membrane Ruffles: Composition, Function, Formation and Visualization.Int J Mol Sci. 2024 Oct 12;25(20):10971. doi: 10.3390/ijms252010971. Int J Mol Sci. 2024. PMID: 39456754 Free PMC article. Review.
-
STIM1 Controls the Focal Adhesion Dynamics and Cell Migration by Regulating SOCE in Osteosarcoma.Int J Mol Sci. 2021 Dec 23;23(1):162. doi: 10.3390/ijms23010162. Int J Mol Sci. 2021. PMID: 35008585 Free PMC article.
References
-
- Sabe H. Requirement for Arf6 in cell adhesion, migration, and cancer cell invasion. J Biochem (Tokyo) 2003;134(4):485–489. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
