

Connecting muscles to tendons: tendons and musculoskeletal development in flies and vertebrates
- PMID: 20699295
- PMCID: PMC2938915
- DOI: 10.1242/dev.047498
Connecting muscles to tendons: tendons and musculoskeletal development in flies and vertebrates
Erratum in
- Development. 2010 Oct;137(19):3347
Abstract
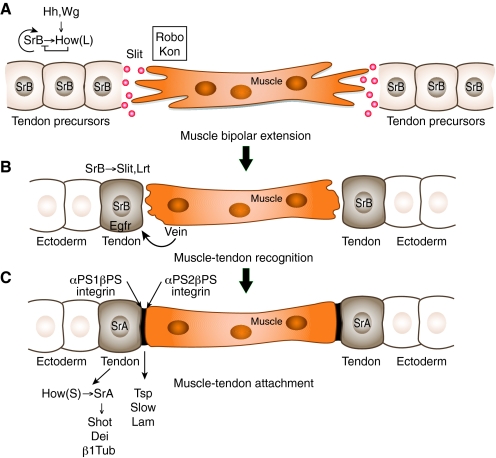
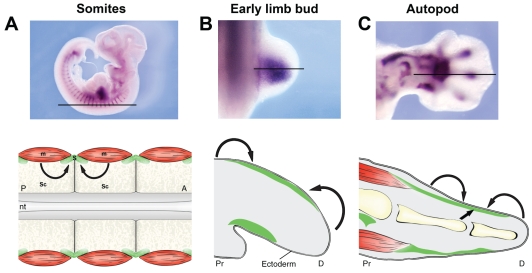
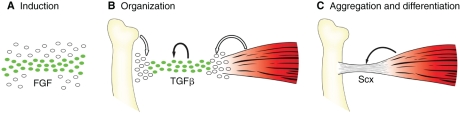
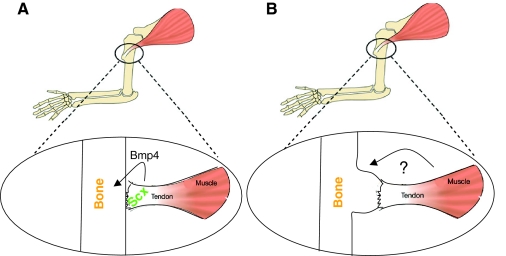
The formation of the musculoskeletal system represents an intricate process of tissue assembly involving heterotypic inductive interactions between tendons, muscles and cartilage. An essential component of all musculoskeletal systems is the anchoring of the force-generating muscles to the solid support of the organism: the skeleton in vertebrates and the exoskeleton in invertebrates. Here, we discuss recent findings that illuminate musculoskeletal assembly in the vertebrate embryo, findings that emphasize the reciprocal interactions between the forming tendons, muscle and cartilage tissues. We also compare these events with those of the corresponding system in the Drosophila embryo, highlighting distinct and common pathways that promote efficient locomotion while preserving the form of the organism.
Figures
Similar articles
-
Development, repair, and regeneration of the limb musculoskeletal system.Curr Top Dev Biol. 2019;132:451-486. doi: 10.1016/bs.ctdb.2018.12.011. Epub 2019 Jan 11. Curr Top Dev Biol. 2019. PMID: 30797517 Review.
-
A non-signaling role of Robo2 in tendons is essential for Slit processing and muscle patterning.Development. 2015 Oct 15;142(20):3512-8. doi: 10.1242/dev.128157. Epub 2015 Sep 23. Development. 2015. PMID: 26400093
-
Building functional units of movement-generation and movement-sensation in the embryo.Int J Dev Biol. 2017;61(3-4-5):171-178. doi: 10.1387/ijdb.160279as. Int J Dev Biol. 2017. PMID: 28621415 Review.
-
Slowdown promotes muscle integrity by modulating integrin-mediated adhesion at the myotendinous junction.Development. 2010 Mar;137(5):785-94. doi: 10.1242/dev.043703. Epub 2010 Jan 28. Development. 2010. PMID: 20110313
-
Coordinated Development of Muscles and Tendon-Like Structures: Early Interactions in the Drosophila Leg.Front Physiol. 2016 Feb 4;7:22. doi: 10.3389/fphys.2016.00022. eCollection 2016. Front Physiol. 2016. PMID: 26869938 Free PMC article.
Cited by
-
Mapping the molecular and structural specialization of the skin basement membrane for inter-tissue interactions.Nat Commun. 2021 May 10;12(1):2577. doi: 10.1038/s41467-021-22881-y. Nat Commun. 2021. PMID: 33972551 Free PMC article.
-
WNT5 interacts with the Ryk receptors doughnut and derailed to mediate muscle attachment site selection in Drosophila melanogaster.PLoS One. 2012;7(3):e32297. doi: 10.1371/journal.pone.0032297. Epub 2012 Mar 5. PLoS One. 2012. PMID: 22403643 Free PMC article.
-
The role of mechanical loading in tendon development, maintenance, injury, and repair.J Bone Joint Surg Am. 2013 Sep 4;95(17):1620-8. doi: 10.2106/JBJS.L.01004. J Bone Joint Surg Am. 2013. PMID: 24005204 Free PMC article. Review.
-
Thrombospondin-4 controls matrix assembly during development and repair of myotendinous junctions.Elife. 2014 Jun 18;3:e02372. doi: 10.7554/eLife.02372. Elife. 2014. PMID: 24941943 Free PMC article.
-
The developmental origin of zygodactyl feet and its possible loss in the evolution of Passeriformes.Proc Biol Sci. 2014 Aug 7;281(1788):20140765. doi: 10.1098/rspb.2014.0765. Proc Biol Sci. 2014. PMID: 24966313 Free PMC article.
References
-
- Akiyama H., Kamitani T., Yang X., Kandyil R., Bridgewater L. C., Fellous M., Mori-Akiyama Y., de Crombrugghe B. (2005). The transcription factor Sox9 is degraded by the ubiquitin-proteasome system and stabilized by a mutation in a ubiquitin-target site. Matrix Biol. 23, 499-505 - PubMed
-
- Becker S., Pasca G., Strumpf D., Min L., Volk T. (1997). Reciprocal signaling between Drosophila epidermal muscle attachment cells and their corresponding muscles. Development 124, 2615-2622 - PubMed
-
- Benjamin M., Ralphs J. R. (1997). Tendons and ligaments – an overview. Histol. Histopathol. 12, 1135-1144 - PubMed
-
- Benjamin M., Kumai T., Milz S., Boszczyk B. M., Boszczyk A. A., Ralphs J. R. (2002). The skeletal attachment of tendons-tendon `entheses'. Comp. Biochem. Physiol. A Physiol. 133, 931-945 - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
