

CHMP2B mutants linked to frontotemporal dementia impair maturation of dendritic spines
- PMID: 20699355
- PMCID: PMC3013364
- DOI: 10.1242/jcs.068817
CHMP2B mutants linked to frontotemporal dementia impair maturation of dendritic spines
Abstract
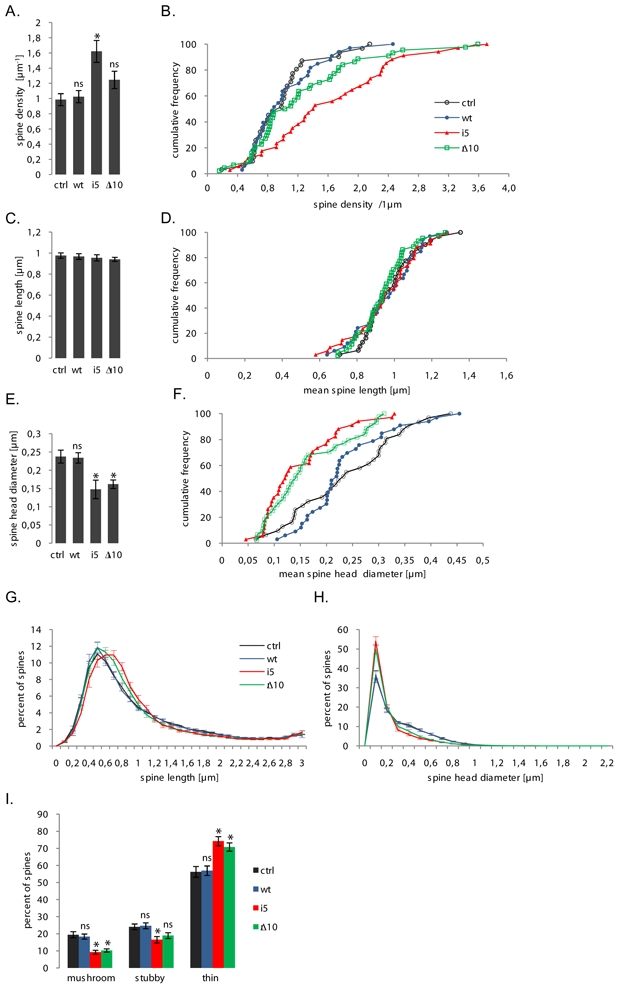
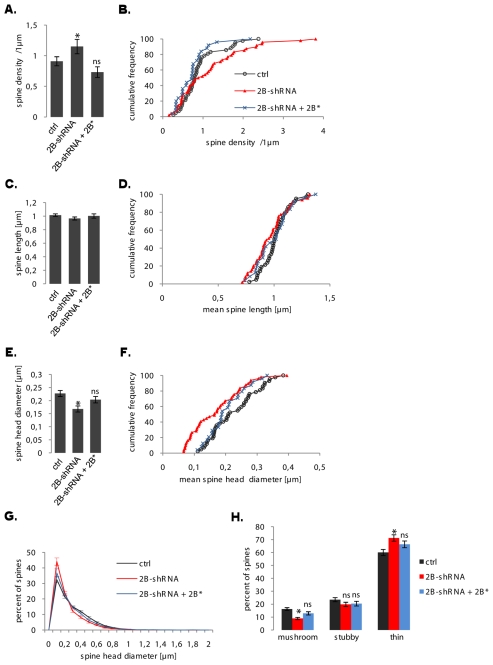
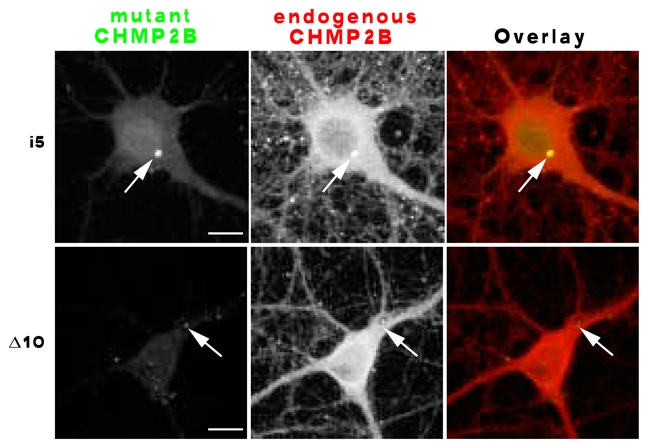
The highly conserved ESCRT-III complex is responsible for deformation and cleavage of membranes during endosomal trafficking and other cellular activities. In humans, dominant mutations in the ESCRT-III subunit CHMP2B cause frontotemporal dementia (FTD). The decade-long process leading to this cortical degeneration is not well understood. One possibility is that, akin to other neurodegenerative diseases, the pathogenic protein affects the integrity of dendritic spines and synapses before any neuronal death. Using confocal microscopy and 3D reconstruction, we examined whether expressing the FTD-linked mutants CHMP2B(intron5) and CHMP2B(Delta10) in cultured hippocampal neurons modified the number or structure of spines. Both mutants induced a significant decrease in the proportion of large spines with mushroom morphology, without overt degeneration. Furthermore, CHMP2B(Delta10) induced a drop in frequency and amplitude of spontaneous excitatory postsynaptic currents, suggesting that the more potent synapses were lost. These effects seemed unrelated to changes in autophagy. Depletion of endogenous CHMP2B by RNAi resulted in morphological changes similar to those induced by mutant CHMP2B, consistent with dominant-negative activity of pathogenic mutants. Thus, CHMP2B is required for spine growth. Taken together, these results demonstrate that a mutant ESCRT-III subunit linked to a human neurodegenerative disease can disrupt the normal pattern of spine development.
Figures







References
-
- Banker G, Goslin K. MIT Press; 1998.
-
- Carlton JG, Martin-Serrano J. Parallels between cytokinesis and retroviral budding: a role for the ESCRT machinery. Science. 2007;316:1908–12. - PubMed
