

The glabra1 mutation affects cuticle formation and plant responses to microbes
- PMID: 20699396
- PMCID: PMC2949009
- DOI: 10.1104/pp.110.161646
The glabra1 mutation affects cuticle formation and plant responses to microbes
Abstract
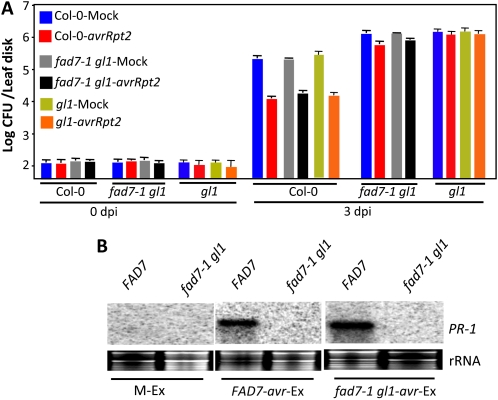
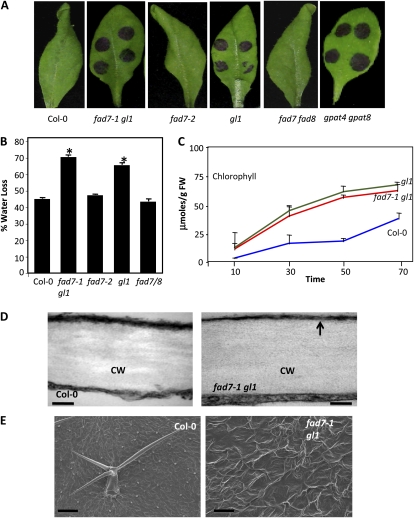
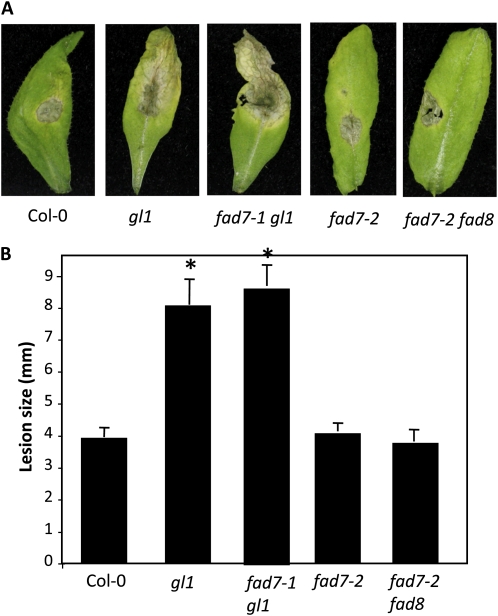
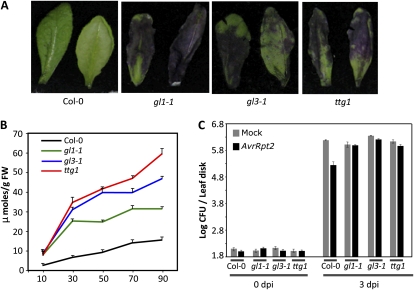
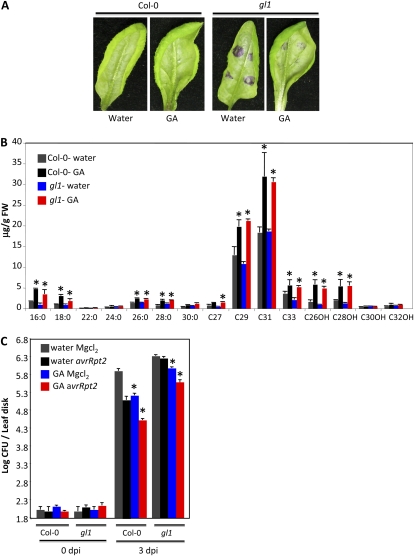
Systemic acquired resistance (SAR) is a form of defense that provides resistance against a broad spectrum of pathogens in plants. Previous work indicates a role for plastidial glycerolipid biosynthesis in SAR. Specifically, mutations in FATTY ACID DESATURASE7 (FAD7), which lead to reduced trienoic fatty acid levels and compromised plastidial lipid biosynthesis, have been associated with defective SAR. We show that the defective SAR in Arabidopsis (Arabidopsis thaliana) fad7-1 plants is not associated with a mutation in FAD7 but rather with a second-site mutation in GLABRA1 (GL1), a gene well known for its role in trichome formation. The compromised SAR in gl1 plants is associated with impairment in their cuticles. Furthermore, mutations in two other components of trichome development, GL3 and TRANSPARENT TESTA GLABRA1, also impaired cuticle development and SAR. This suggests an overlap in the biochemical pathways leading to cuticle and trichome development. Interestingly, exogenous application of gibberellic acid (GA) not only enhanced SAR in wild-type plants but also restored SAR in gl1 plants. In contrast to GA, the defense phytohoromes salicylic acid and jasmonic acid were unable to restore SAR in gl1 plants. GA application increased levels of cuticular components but not trichome formation on gl1 plants, thus implicating cuticle, but not trichomes, as an important component of SAR. Our findings question the prudence of using mutant backgrounds for genetic screens and underscore a need to reevaluate phenotypes previously studied in the gl1 background.
Figures
References
-
- Achard P, Genschik P. (2009) Releasing the brakes of plant growth: how GAs shutdown DELLA proteins. J Exp Bot 60: 1085–1092 - PubMed
-
- Achard P, Renou JP, Berthome R, Harberd NP, Genschik P. (2008) Plant DELLAs restrain growth and promote survival of adversity by reducing the levels of reactive oxygen species. Curr Biol 18: 656–660 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
