

DsbA-L alleviates endoplasmic reticulum stress-induced adiponectin downregulation
- PMID: 20699416
- PMCID: PMC2963539
- DOI: 10.2337/db10-0412
DsbA-L alleviates endoplasmic reticulum stress-induced adiponectin downregulation
Abstract
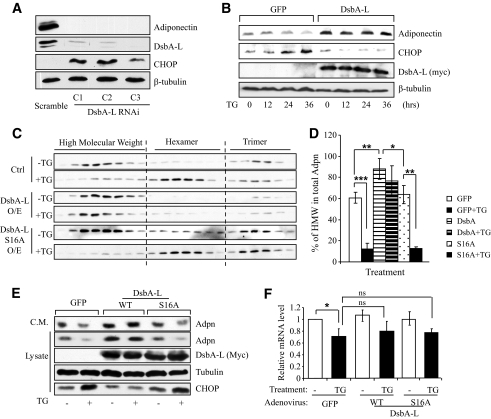
Objective: Obesity impairs adiponectin expression, assembly, and secretion, yet the underlying mechanisms remain elusive. The aims of this study were 1) to determine the molecular mechanisms by which obesity impairs adiponectin multimerization and stability, and 2) to determine the potential role of disulfide-bond-A oxidoreductase-like protein (DsbA-L), a recently identified adiponectin interactive protein that promotes adiponectin multimerization and stability in obesity-induced endoplasmic reticulum (ER) stress and adiponectin downregulation.
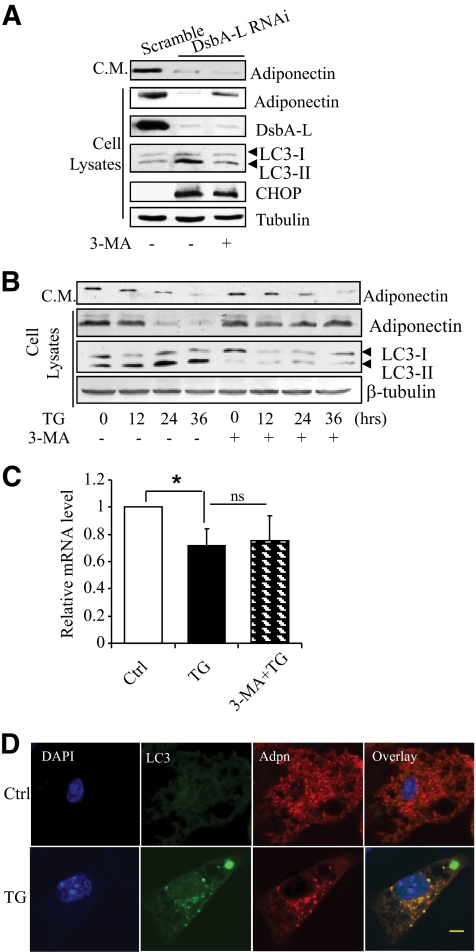
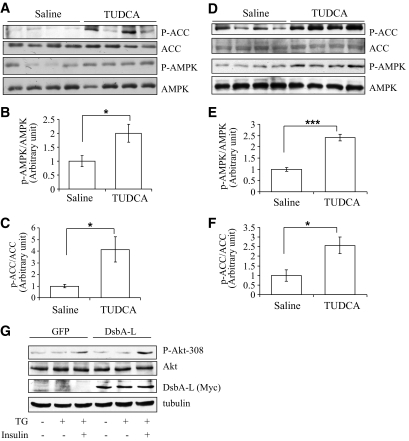
Research design and methods: Tauroursodeoxycholic acid (TUDCA), a chemical chaperone that alleviates ER stress, was used to study the mechanism underlying obesity-induced adiponectin downregulation in db/db mice, high-fat diet-induced obese mice, and in ER-stressed 3T3-L1 adipocytes. The cellular levels of DsbA-L were altered by RNAi-mediated suppression or adenovirus-mediated overexpression. The protective role of DsbA-L in obesity- and ER stress-induced adiponectin downregulation was characterized.
Results: Treating db/db mice and diet-induced obese mice with TUDCA increased the cellular and serum levels of adiponectin. In addition, inducing ER stress is sufficient to downregulate adiponectin levels in 3T3-L1 adipocytes, which could be protected by treating cells with the autophagy inhibitor 3-methyladenine or by overexpression of DsbA-L.
Conclusions: ER stress plays a key role in obesity-induced adiponectin downregulation. In addition, DsbA-L facilitates adiponectin folding and assembly and provides a protective effect against ER stress-mediated adiponectin downregulation in obesity.
Figures

Similar articles
-
Endoplasmic reticulum (ER) localization is critical for DsbA-L protein to suppress ER stress and adiponectin down-regulation in adipocytes.J Biol Chem. 2015 Apr 17;290(16):10143-8. doi: 10.1074/jbc.M115.645416. Epub 2015 Mar 4. J Biol Chem. 2015. PMID: 25739441 Free PMC article.
-
Autophagy: roles in obesity-induced ER stress and adiponectin downregulation in adipocytes.Autophagy. 2010 Nov;6(8):1196-7. doi: 10.4161/auto.6.8.13478. Epub 2010 Nov 16. Autophagy. 2010. PMID: 20864818 Free PMC article.
-
Autophagy-mediated insulin receptor down-regulation contributes to endoplasmic reticulum stress-induced insulin resistance.Mol Pharmacol. 2009 Sep;76(3):596-603. doi: 10.1124/mol.109.057067. Epub 2009 Jun 18. Mol Pharmacol. 2009. PMID: 19541767 Free PMC article.
-
[A Multifaceted Approach regarding the Association of the DsbA-L Gene with the Risk of Obesity-related Diseases Based on Clinical Pharmacogenetics].Yakugaku Zasshi. 2019;139(1):53-60. doi: 10.1248/yakushi.18-00163-3. Yakugaku Zasshi. 2019. PMID: 30606929 Review. Japanese.
-
Regulation of adiponectin multimerization, signaling and function.Best Pract Res Clin Endocrinol Metab. 2014 Jan;28(1):25-31. doi: 10.1016/j.beem.2013.06.003. Epub 2013 Jul 5. Best Pract Res Clin Endocrinol Metab. 2014. PMID: 24417943 Free PMC article. Review.
Cited by
-
Synthetic peptides designed to modulate adiponectin assembly improve obesity-related metabolic disorders.Br J Pharmacol. 2017 Dec;174(23):4478-4492. doi: 10.1111/bph.14050. Epub 2017 Nov 2. Br J Pharmacol. 2017. PMID: 28945274 Free PMC article.
-
The cGAS-STING signaling in cardiovascular and metabolic diseases: Future novel target option for pharmacotherapy.Acta Pharm Sin B. 2022 Jan;12(1):50-75. doi: 10.1016/j.apsb.2021.05.011. Epub 2021 May 20. Acta Pharm Sin B. 2022. PMID: 35127372 Free PMC article. Review.
-
Adipocyte spliced form of X-box-binding protein 1 promotes adiponectin multimerization and systemic glucose homeostasis.Diabetes. 2014 Mar;63(3):867-79. doi: 10.2337/db13-1067. Epub 2013 Nov 15. Diabetes. 2014. PMID: 24241534 Free PMC article.
-
Adiponectin for the treatment of diabetic nephropathy.Korean J Intern Med. 2019 May;34(3):480-491. doi: 10.3904/kjim.2019.109. Epub 2019 Apr 30. Korean J Intern Med. 2019. PMID: 31048658 Free PMC article. Review.
-
DsbA-L prevents obesity-induced inflammation and insulin resistance by suppressing the mtDNA release-activated cGAS-cGAMP-STING pathway.Proc Natl Acad Sci U S A. 2017 Nov 14;114(46):12196-12201. doi: 10.1073/pnas.1708744114. Epub 2017 Oct 30. Proc Natl Acad Sci U S A. 2017. PMID: 29087318 Free PMC article.
References
-
- Shetty S, Kusminski CM, Scherer PE. Adiponectin in health and disease: evaluation of adiponectin-targeted drug development strategies. Trends Pharmacol Sci 2009;30:234–239 - PubMed
-
- Pajvani UB, Du X, Combs TP, Berg AH, Rajala MW, Schulthess T, Engel J, Brownlee M, Scherer PE. Structure-function studies of the adipocyte-secreted hormone Acrp30/adiponectin. Implications fpr metabolic regulation and bioactivity. J Biol Chem 2003;278:9073–9085 - PubMed
-
- Tsao TS, Murrey HE, Hug C, Lee DH, Lodish HF. Oligomerization state-dependent activation of NF-κ B signaling pathway by adipocyte complement-related protein of 30 kDa (Acrp30). J Biol Chem 2002;277:29359–29362 - PubMed
-
- Waki H, Yamauchi T, Kamon J, Ito Y, Uchida S, Kita S, Hara K, Hada Y, Vasseur F, Froguel P, Kimura S, Nagai R, Kadowaki T. Impaired multimerization of human adiponectin mutants associated with diabetes. Molecular structure and multimer formation of adiponectin. J Biol Chem 2003;278:40352–40363 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
