

Central mechanisms of odour object perception
- PMID: 20700142
- PMCID: PMC3722866
- DOI: 10.1038/nrn2883
Central mechanisms of odour object perception
Abstract
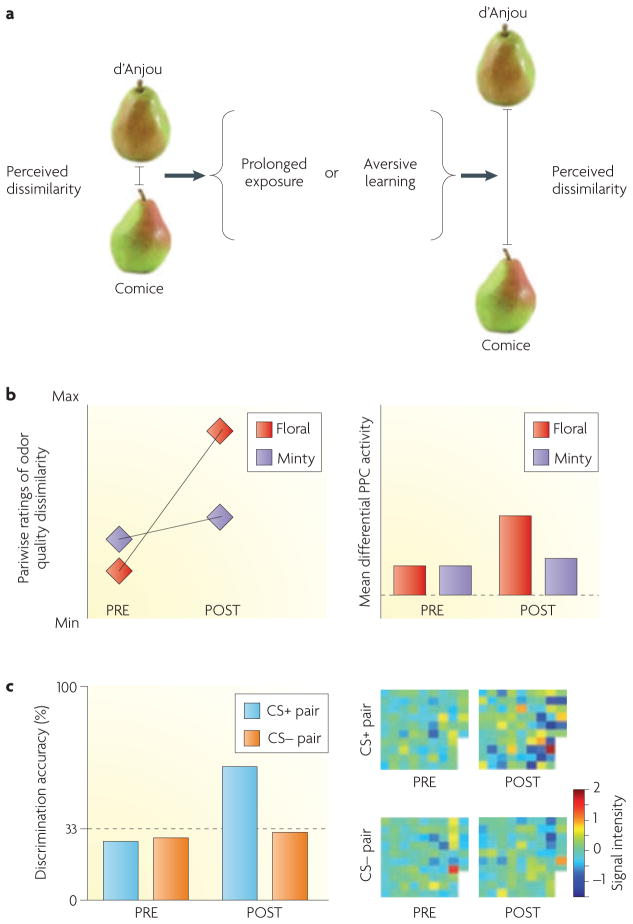
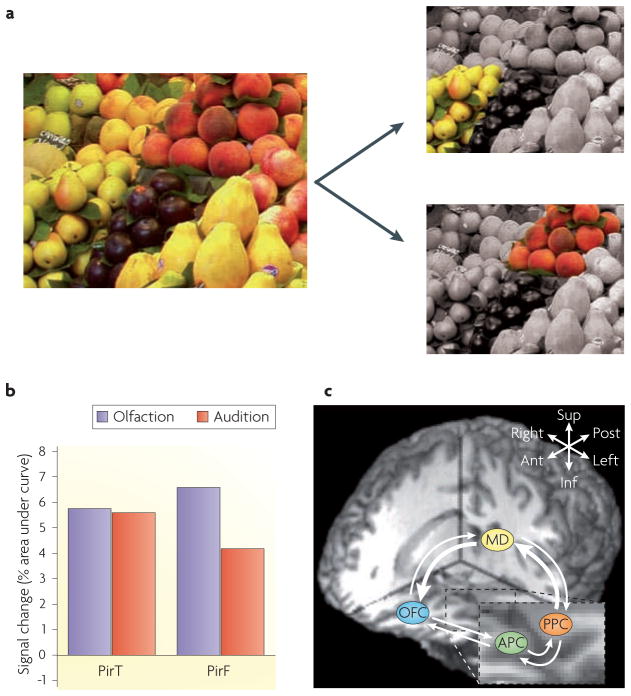
The stimulus complexity of naturally occurring odours presents unique challenges for central nervous systems that are aiming to internalize the external olfactory landscape. One mechanism by which the brain encodes perceptual representations of behaviourally relevant smells is through the synthesis of different olfactory inputs into a unified perceptual experience--an odour object. Recent evidence indicates that the identification, categorization and discrimination of olfactory stimuli rely on the formation and modulation of odour objects in the piriform cortex. Convergent findings from human and rodent models suggest that distributed piriform ensemble patterns of olfactory qualities and categories are crucial for maintaining the perceptual constancy of ecologically inconstant stimuli.
Conflict of interest statement
The author declares no competing financial interests.
Figures
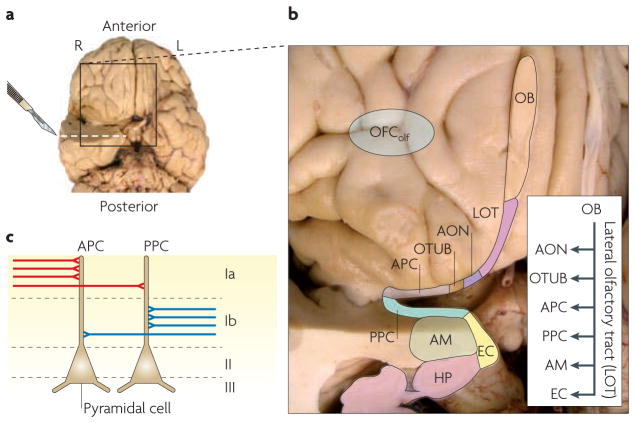
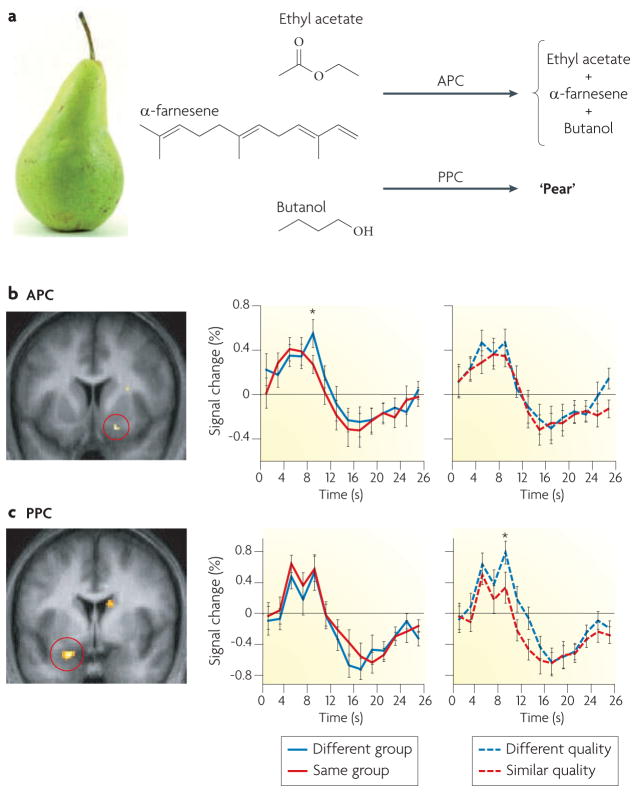
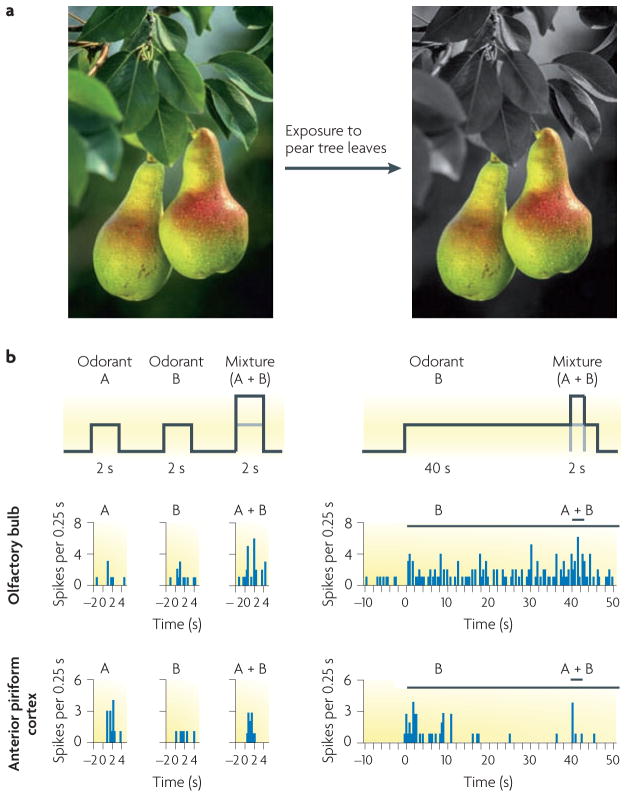
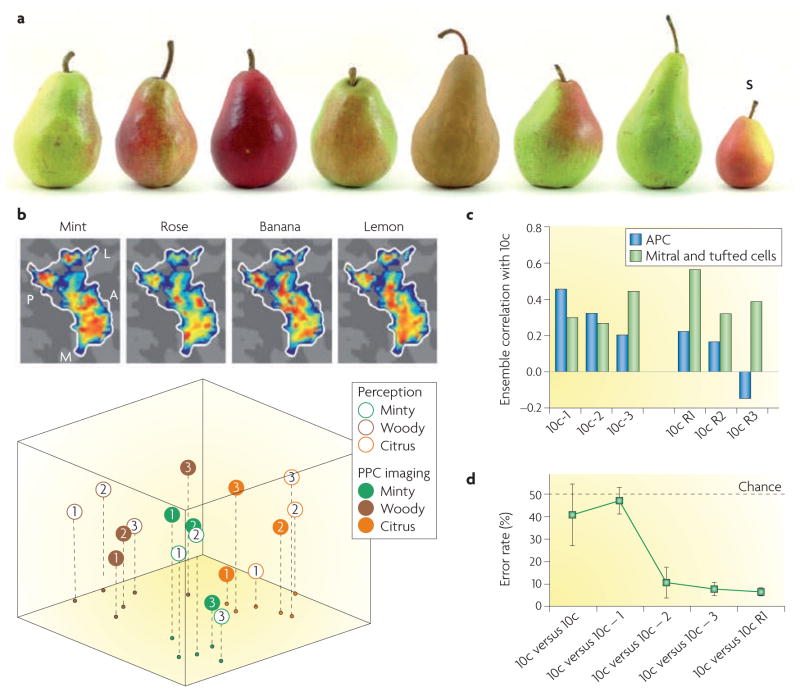
References
-
- Aristotle. On Sense and the Sensible. The Internet Classics Archive. 350 BC [online], http://classics.mit.edu/Aristotle/sense.html.
-
- Gross CG. Representation of visual stimuli in inferior temporal cortex. Phil Trans R Soc Lond B. 1992;335:3–10. - PubMed
-
- Ungerleider LG, Haxby JV. ‘What’ and ‘where’ in the human brain. Curr Opin Neurobiol. 1994;4:157–165. - PubMed
-
- Logothetis NK, Sheinberg DL. Visual object recognition. Annu Rev Neurosci. 1996;19:577–621. - PubMed
-
- Tanaka K. Inferotemporal cortex and object vision. Annu Rev Neurosci. 1996;19:109–139. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
