

How cofilin severs an actin filament
- PMID: 20700473
- PMCID: PMC2917815
- DOI: 10.1007/s12551-009-0008-5
How cofilin severs an actin filament
Abstract
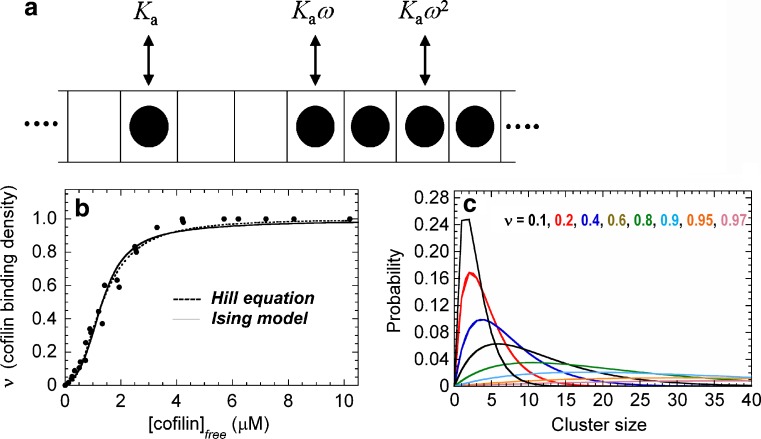
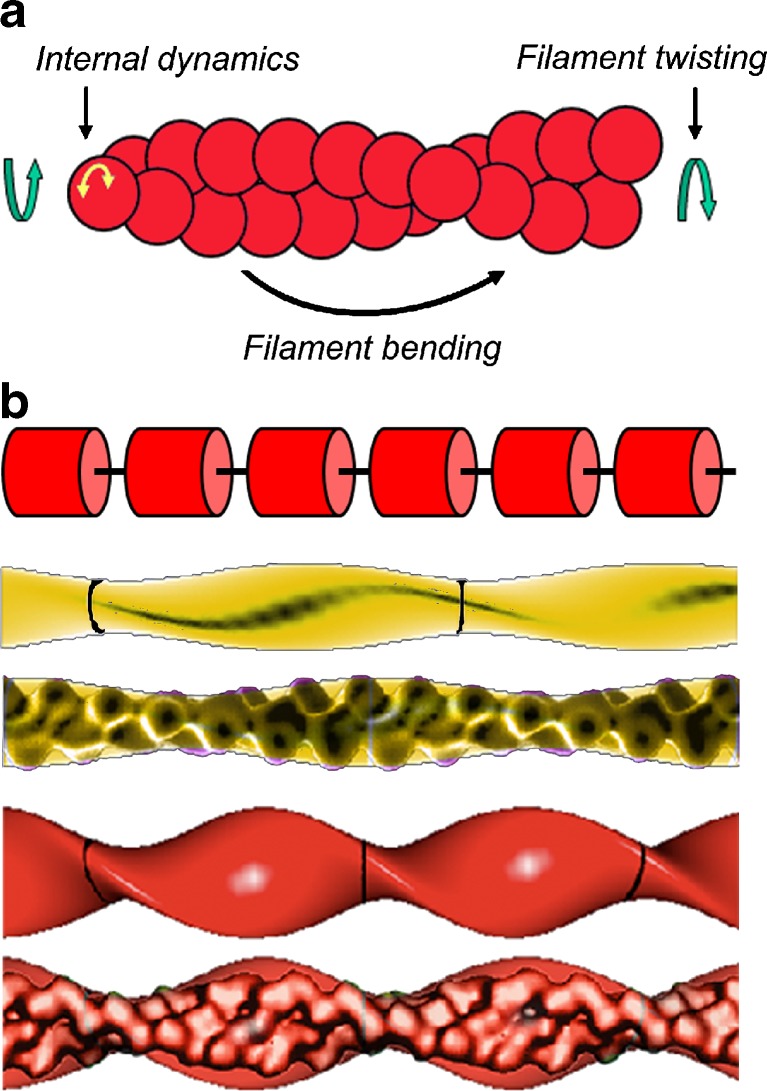
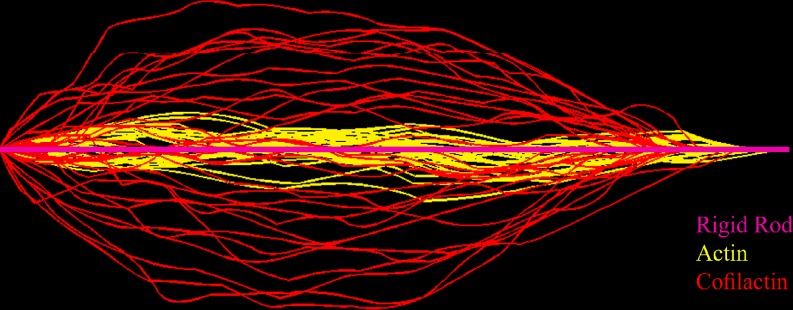
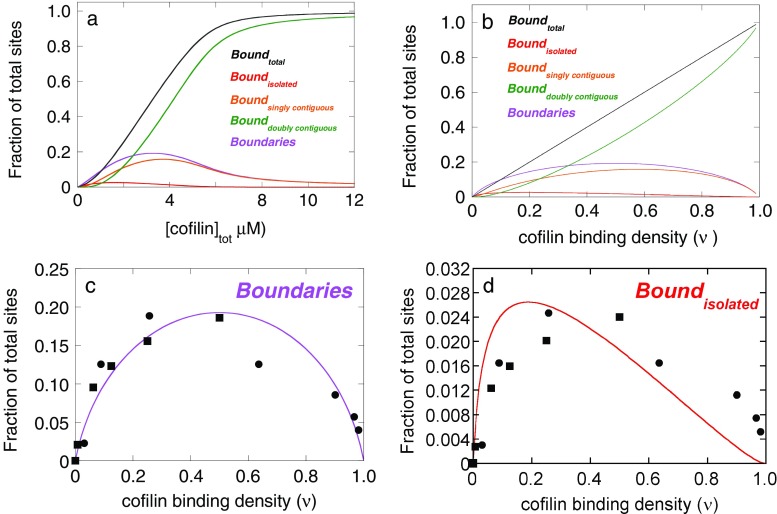
The actin regulatory protein, cofilin, promotes actin assembly dynamics by severing filaments and increasing the number of ends from which subunits add and dissociate. Recent studies provide biophysical descriptions of cooperative filament interactions in energetic, mechanical and structural terms. A one-dimensional Ising model with nearest-neighbor interactions permits thermodynamic analysis of cooperative binding and indicates that one or a few cofilin molecules can sever a filament. Binding and cooperative interactions are entropically driven. A significant fraction of the binding free energy results from the linked dissociation of filament-associated ions (polyelectrolyte effect), which modulate filament structure, stability and mechanics. The remaining binding free energy and essentially all of the cooperative free energy arise from the enhanced conformational dynamics of the cofilactin complex. Filament mechanics are modulated by cofilin such that cofilin-saturated filaments are approximately 10- to 20-fold more compliant in bending and twisting than bare filaments. Cofilin activity is well described by models in which discontinuities in topology, mechanics and conformational dynamics generate stress concentration and promote fracture at junctions of bare and decorated segments, analogous to the grain boundary fracture of crystalline materials and the thermally driven formation of shear transformation zones in colloidal glass.
Figures
Similar articles
-
Cofilin-linked changes in actin filament flexibility promote severing.Biophys J. 2011 Jul 6;101(1):151-9. doi: 10.1016/j.bpj.2011.05.049. Biophys J. 2011. PMID: 21723825 Free PMC article.
-
Clusters of a Few Bound Cofilins Sever Actin Filaments.J Mol Biol. 2021 Apr 2;433(7):166833. doi: 10.1016/j.jmb.2021.166833. Epub 2021 Jan 30. J Mol Biol. 2021. PMID: 33524412 Free PMC article.
-
The kinetics of cooperative cofilin binding reveals two states of the cofilin-actin filament.Biophys J. 2010 May 19;98(9):1893-901. doi: 10.1016/j.bpj.2010.01.023. Biophys J. 2010. PMID: 20441753 Free PMC article.
-
Biophysics of actin filament severing by cofilin.FEBS Lett. 2013 Apr 17;587(8):1215-9. doi: 10.1016/j.febslet.2013.01.062. Epub 2013 Feb 5. FEBS Lett. 2013. PMID: 23395798 Free PMC article. Review.
-
Regulation of actin filament dynamics by actin depolymerizing factor/cofilin and actin-interacting protein 1: new blades for twisted filaments.Biochemistry. 2003 Nov 25;42(46):13363-70. doi: 10.1021/bi034600x. Biochemistry. 2003. PMID: 14621980 Review.
Cited by
-
Arabidopsis CDPK6 phosphorylates ADF1 at N-terminal serine 6 predominantly.Plant Cell Rep. 2013 Nov;32(11):1715-28. doi: 10.1007/s00299-013-1482-6. Epub 2013 Aug 1. Plant Cell Rep. 2013. PMID: 23903947
-
Torsional stress generated by ADF/cofilin on cross-linked actin filaments boosts their severing.Proc Natl Acad Sci U S A. 2019 Feb 12;116(7):2595-2602. doi: 10.1073/pnas.1812053116. Epub 2019 Jan 28. Proc Natl Acad Sci U S A. 2019. PMID: 30692249 Free PMC article.
-
INF2-mediated severing through actin filament encirclement and disruption.Curr Biol. 2014 Jan 20;24(2):156-164. doi: 10.1016/j.cub.2013.12.018. Epub 2014 Jan 9. Curr Biol. 2014. PMID: 24412206 Free PMC article.
-
Origin of twist-bend coupling in actin filaments.Biophys J. 2010 Sep 22;99(6):1852-60. doi: 10.1016/j.bpj.2010.07.009. Biophys J. 2010. PMID: 20858430 Free PMC article.
-
Cofilin-induced unidirectional cooperative conformational changes in actin filaments revealed by high-speed atomic force microscopy.Elife. 2015 Feb 2;4:e04806. doi: 10.7554/eLife.04806. Elife. 2015. PMID: 25642645 Free PMC article.
References
-
- Ackbarow T, Buehler MJ. Superelasticity, energy dissipation and strain hardening of vimentin coiled-coil intermediate filaments. J Mater Sci. 2007;21:2855–2869.
-
- Anderson TL. Fracture Mechanics: Fundamentals and Applications, 3 rd edit. Boca Raton, FL: CRC Press; 2005.
-
- Barkley MD, Zimm BH. Theory of twisting and bending of chain molecules - Analysis of the fluorescence depolarization of DNA. J Chem Phys. 1979;70:2991–3007. doi: 10.1063/1.437838. - DOI
Grants and funding
LinkOut - more resources
Full Text Sources
