

Alternative processing of scrambled genes generates protein diversity in the ciliate Chilodonella uncinata
- PMID: 20700892
- PMCID: PMC2933047
- DOI: 10.1002/jez.b.21354
Alternative processing of scrambled genes generates protein diversity in the ciliate Chilodonella uncinata
Abstract
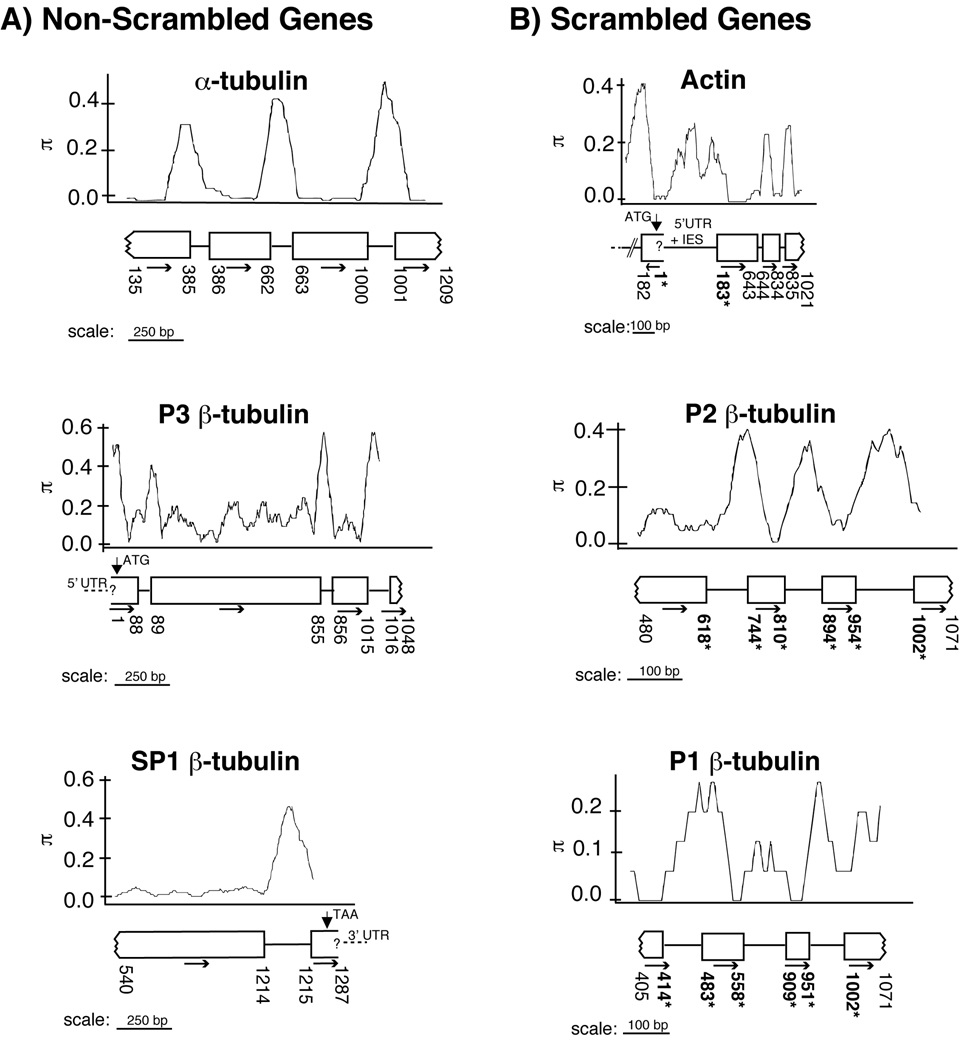
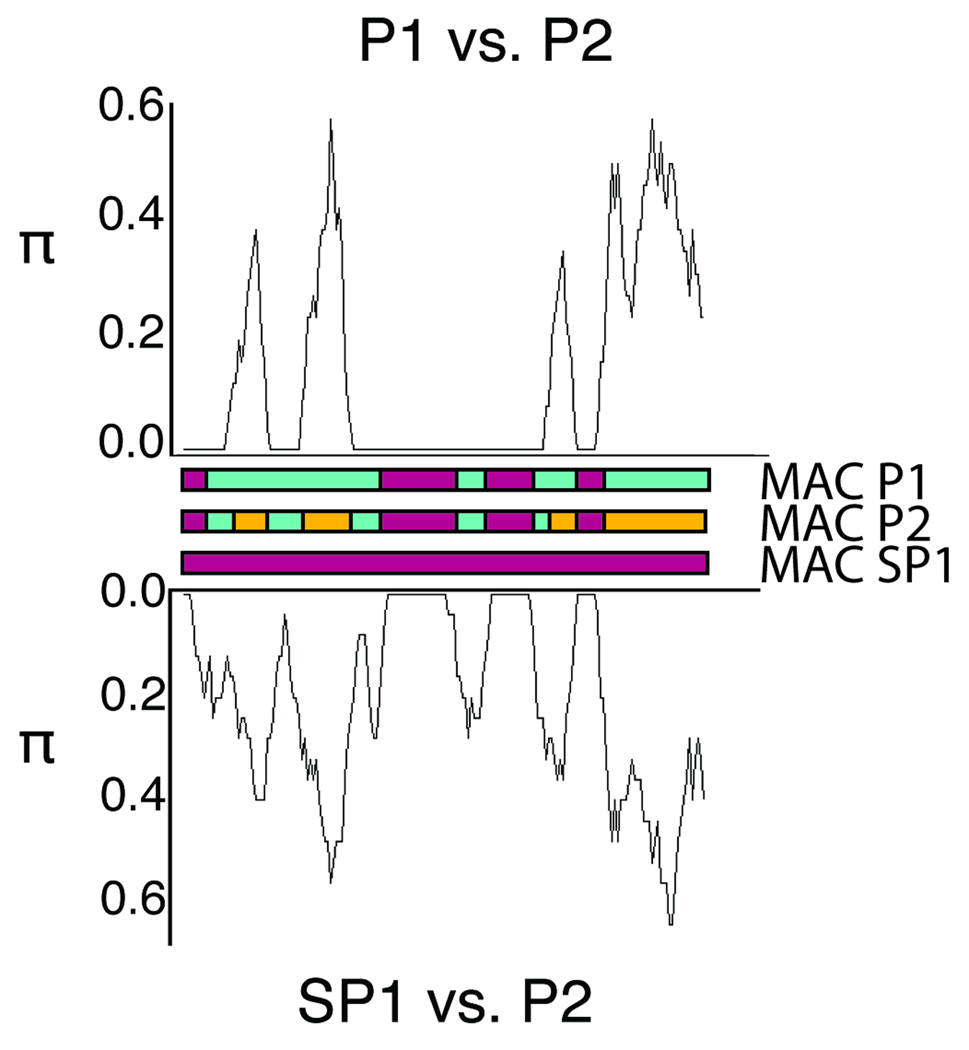
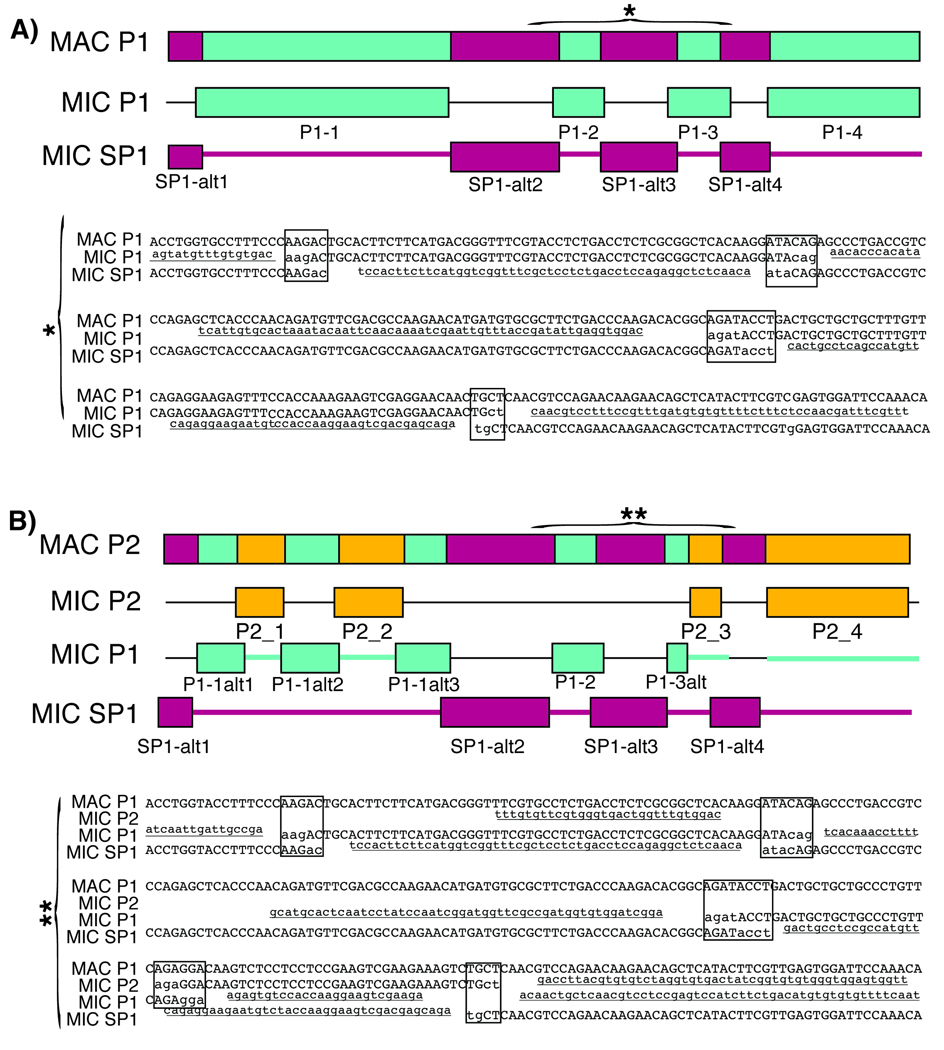
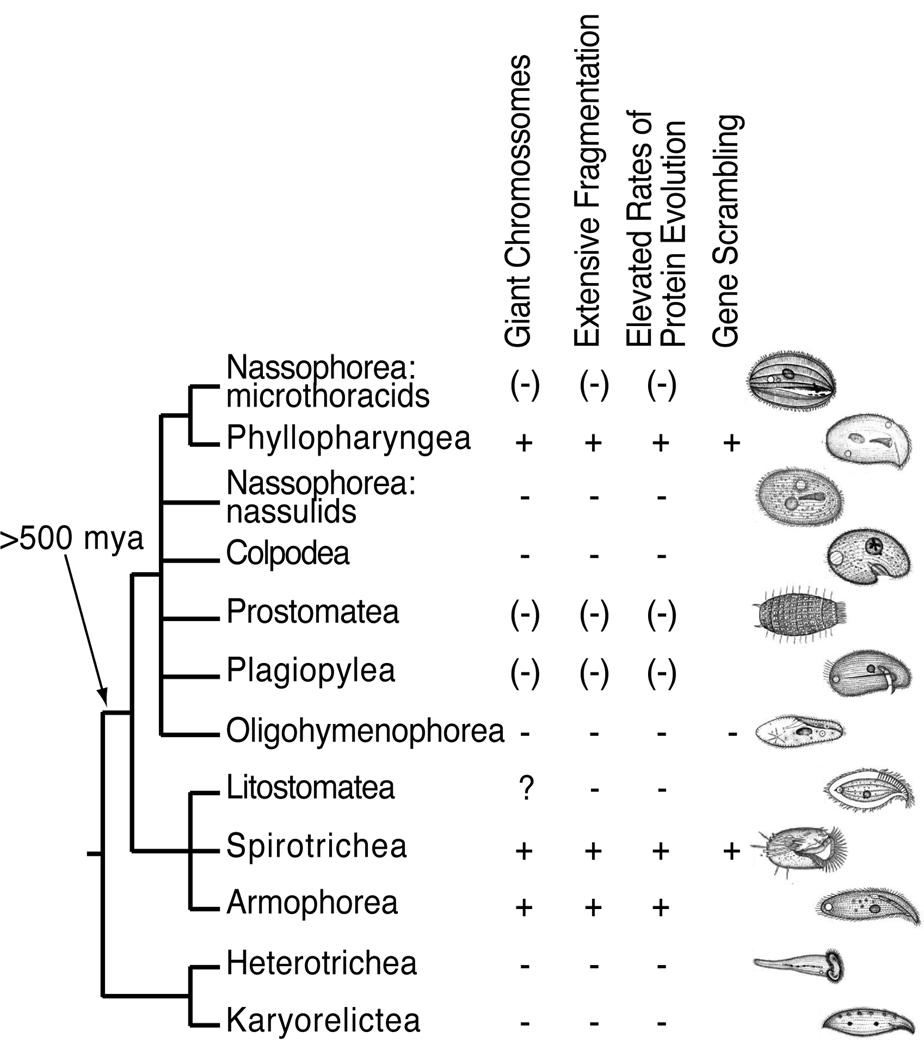
In ciliates, chromosomal rearrangements occur during the development of the somatic macronuclear genome from the germline micronuclear genome. These rearrangements are extensive in three ciliate classes-Armophorea, Spirotrichea, and Phyllopharyngea-generating a macronucleus with up to 20,000,000 gene-sized chromosomes. Earlier, we have shown that these three classes also share elevated rates of protein evolution relative to other ciliates. To assess the evolution of germline-limited sequences in the class Phyllopharyngea, we used a combination of traditional and walking PCR to analyze micronuclear copies of multiple genes from two lines of the morphospecies Chilodonella uncinata for which we had previously characterized macronuclear sequences. Analyses of the resulting data yield three main results: (1) conserved macronuclear (somatic) regions are found within rapidly evolving micronuclear (germline) regions; (2) gene scrambling exists within this ciliate lineage; and (3) alternative processing of micronuclear regions yields diverse macronuclear beta-tubulin paralogs. To our knowledge, this is the first study to demonstrate gene scrambling outside the nonsister class Spirotrichea, and to show that alternative processing of scrambled genes generates diversity in gene families. Intriguingly, the Spirotrichea and Phyllopharyngea are also united in having transient "giant" polytene chromosomes, gene-sized somatic chromosomes, and elevated rates of protein evolution. We hypothesize that this suite of characters enables these ciliates to enjoy the benefits of asexuality while still maintaining the ability to go through sexual cycles. The data presented here add to the growing evidence of the dynamic nature of eukaryotic genomes within diverse lineages across the tree of life.
(c) 2010 Wiley-Liss, Inc.
Figures
References
-
- Ammermann D. Giant chromosomes in ciliates. Results Probs Cell Differ. 1987;14:59–67. - PubMed
-
- Ausubel FM, Brent R, Kingston RE, Moore DD, Seidman JG, Smith JA, Struhl K. Current Protocols in Molecular Biology. New York: Wiley-Liss; 1993.
-
- Curtis EA, Landweber LF. Evolution of gene scrambling in ciliate micronuclear genes. Ann N Y Acad Sci. 1999;870:349–350. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
