

Regulation of response regulator autophosphorylation through interdomain contacts
- PMID: 20702407
- PMCID: PMC2952233
- DOI: 10.1074/jbc.M110.157164
Regulation of response regulator autophosphorylation through interdomain contacts
Abstract
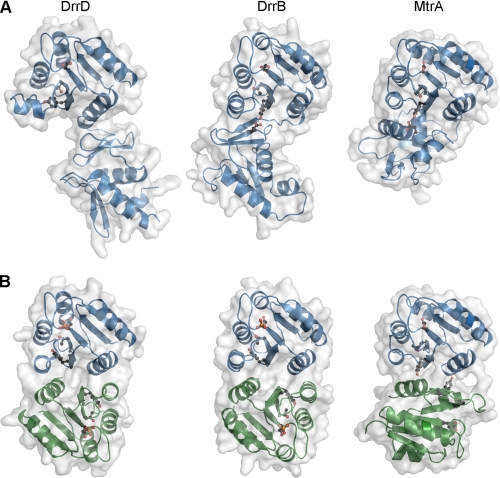
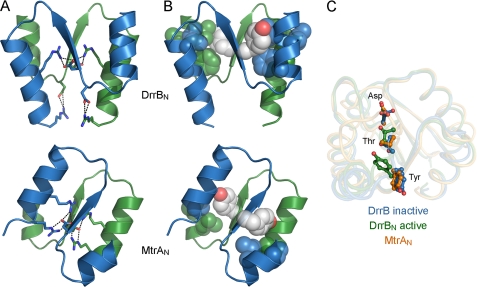
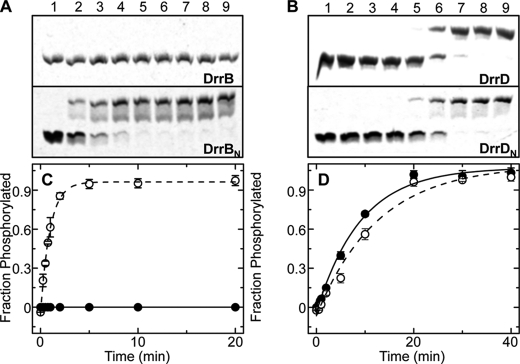
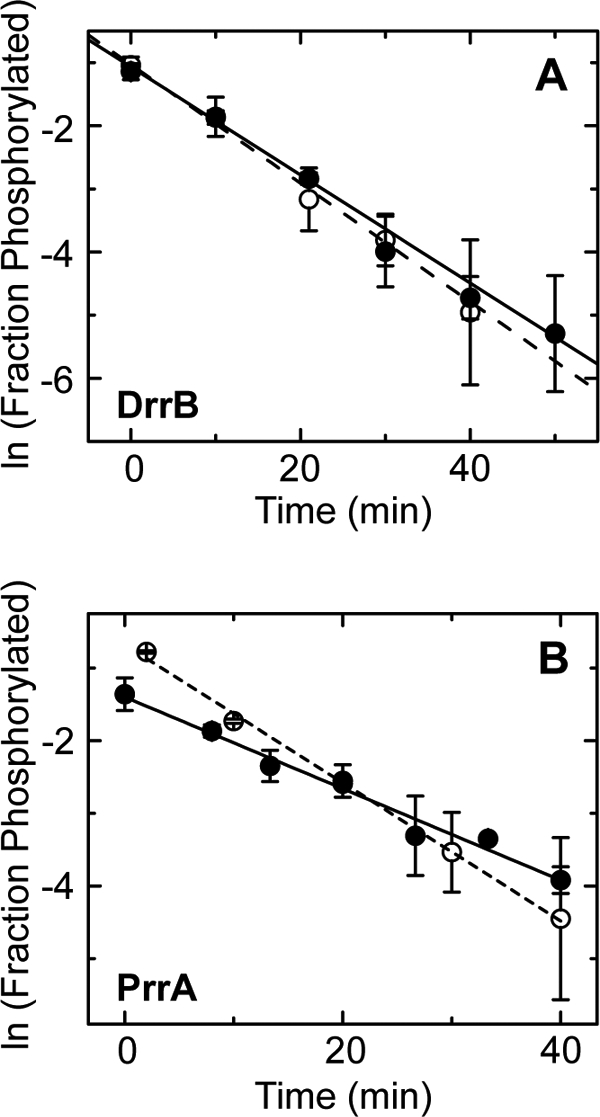
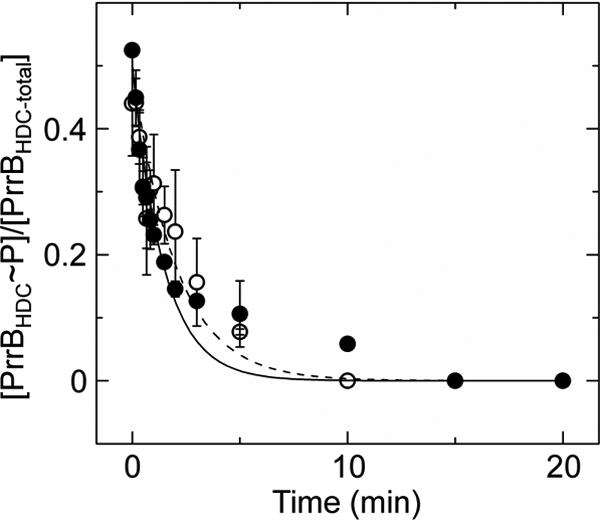
DNA-binding response regulators (RRs) of the OmpR/PhoB subfamily alternate between inactive and active conformational states, with the latter having enhanced DNA-binding affinity. Phosphorylation of an aspartate residue in the receiver domain, usually via phosphotransfer from a cognate histidine kinase, stabilizes the active conformation. Many of the available structures of inactive OmpR/PhoB family proteins exhibit extensive interfaces between the N-terminal receiver and C-terminal DNA-binding domains. These interfaces invariably involve the α4-β5-α5 face of the receiver domain, the locus of the largest differences between inactive and active conformations and the surface that mediates dimerization of receiver domains in the active state. Structures of receiver domain dimers of DrrB, DrrD, and MtrA have been determined, and phosphorylation kinetics were analyzed. Analysis of phosphotransfer from small molecule phosphodonors has revealed large differences in autophosphorylation rates among OmpR/PhoB RRs. RRs with substantial domain interfaces exhibit slow rates of phosphorylation. Rates are greatly increased in isolated receiver domain constructs. Such differences are not observed between autophosphorylation rates of full-length and isolated receiver domains of a RR that lacks interdomain interfaces, and they are not observed in histidine kinase-mediated phosphotransfer. These findings suggest that domain interfaces restrict receiver domain conformational dynamics, stabilizing an inactive conformation that is catalytically incompetent for phosphotransfer from small molecule phosphodonors. Inhibition of phosphotransfer by domain interfaces provides an explanation for the observation that some RRs cannot be phosphorylated by small molecule phosphodonors in vitro and provides a potential mechanism for insulating some RRs from small molecule-mediated phosphorylation in vivo.
Figures
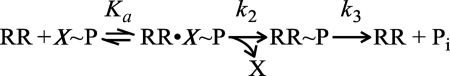

Comment in
-
Influence of interdomain interfaces. Regulation of response regulator autophosphorylation through interdomain contacts.J Biol Chem. 2010 Oct 15;285(42):e99975. doi: 10.1074/jbc.p110.157164. J Biol Chem. 2010. PMID: 20954302 Free PMC article. No abstract available.
References
-
- Hoch J. A., Silhavy T. J. (eds) (1995) Two-component Signal Transduction, American Society for Microbiology Press, Washington, DC
-
- Stock A. M., Robinson V. L., Goudreau P. N. (2000) Annu. Rev. Biochem. 69, 183–215 - PubMed
-
- Volkman B. F., Lipson D., Wemmer D. E., Kern D. (2001) Science 291, 2429–2433 - PubMed
-
- Nohaile M., Kern D., Wemmer D., Stedman K., Kustu S. (1997) J. Mol. Biol. 273, 299–316 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
